Mesozoic Birds
Avialae Gauthier, 1986
The birds originated from the Maniraptora branch of Theropod dinosaurs. Avialae is defined as all theropods closer to modern birds than to Deinonychus. Thus the Deinonychosauria (raptor dinosaurs: dromaeosaurids and troodontids) are not part of Avialae. Other Maniraptora that are not part of Avialae include Oviraptorosauria (egg thief dinosaurs), Therizinosauria (sloth dinosaurs), Ornitholestes (bird robber) and the Alvarezsauroidea (thumb-claw dinosaurs). The Ornithomimosauria (ostrich dinosaurs) are the sister group to Maniraptora. All are rather bird-like, but powered flight seems to have originated in Avialae.
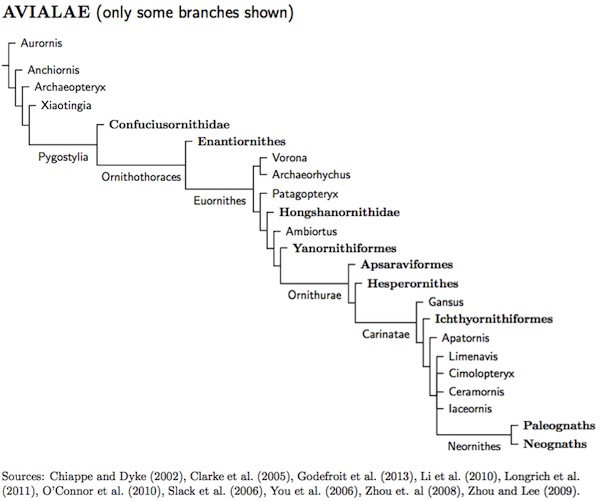
The Urvögel
The Archaeopteryx has been referred to as the urvögel, the first bird, since the mid-19th century. It's known from fossils in the Solnhofen limestone in Bavaria, southern Germany. These fossils are about 145-150 million years old, near the end of the Jurassic period. Recent discoveries have found older (160 million-year-old) bird-like fossils such as Anchiornis, Xiaotingia, and Aurornis that have cast doubt on its status as the first bird (see Godefroit et al., 2013). Moreover, questions have been raised about whether it could fly and whether it is truly in the lineage leading to modern birds, or perhaps in Deinonychosauria instead.
Like other early birds, Archaeopteryx had a long bony tail. However, modern birds have a different arrangement. The tail is composed of feathers attached to a bony structure called a pygostyle, made up of a fused vertebrae. Since the bony tail is short, birds with a pygostyle are sometimes called short-tailed birds, even though they may have very long feather tails (think peacocks).
Pygostylia Chiappe, 1997
Pygostylia contains all the short-tailed birds. Chiappe (1997) defined it as the smallest clade containing both the modern birds and the Confucius birds (Confuciusornithidae). The Pygostylia are thought to have originated early in the Cretaceous period, about 131-140 million years ago. Certaintly they were around 130 million years ago.
Many of the Pygostylia have a long rod or blade for a pygostyle. In the case of the Confuciusornithidae, which a pair of ribbon-like tail feathers were attached to the pygostyle. One early Confucius bird was Eoconfuciusornis, which dates to about 131 million years ago. Numerous fossils of other Confucius birds have come from the Jehol fossil beds in China, with the latest specimens dating to about 120 million years ago.
Ornithothoraces: Enantiornithines and Euornithes Chiappe and Calvo, 1994
The next group on the road to modern is Ornithothoraces. This clade is defined as the smallest clade including both the opposite bird Iberomesornis and modern birds. Thus it contains all Enantiornithines (opposite birds), modern birds, and more.
Enantiornithines Walker, 1981
The Enantiornithines, or opposite birds were only recognized as distinct group in 1981. The Enantiornithines are called opposite birds because the articulation in the shoulder is reversed compared with modern birds. They also have the primitive form of pygostyle. They were quite common at some point, and over 50 species are recognized. The earliest opposite birds date to at least 130 million years ago, and the most recent were present until the end mass extinction at the end of the Cretaceous. Fossils of Avisaurus and other Enantiornithines have been found in the Lance formation in Wyoming, which was laid down in the last years of the Cretaceous. At times, the opposite birds seem to be the dominant portion of the avifauna. However, that was not true at Lance Creek near the end of the Cretaceous.
Euornithes Cope, 1889
Euornithes is almost the same as the slightly less inclusive Ornithuromorpha. It refers to all birds that are more closely related to modern birds than to the opposite bird Sinornis, and hence closer to modern birds than to any of the Enantiornithines (Serrano, 1999). The earliest known fossils (Ambiortus) date to about 130 or more million years ago. Most of the Euornithes have the shovel-shaped modern form of pygostyle.
One notable early branch of the Euornithes is the Yanornithiformes, known from specimens found in China in the Yixian and Jiufotang formations (120-125 million years ago). Another notable branch from the Yixian and Jiufotang formations is the Hongshanornithidae, which may also be part of the Yanornithiformes. Neither group appears to be on the lineage leading to modern birds, and fossils from them are only known from the early Cretaceous.
Ornithurae Haeckel, 1866
One definition of the Ornithurae is that it is the largest clade containing Ichthyornis, Hesperornis, and the modern birds. Other definitions are more expansive and include the closely related Apsaraviformes (Kurochkin, 1982).
Two Apsaraviformes are known, Apsaravis and Palintropus. Palintropus had once been considered a Neonithe, but it isn't. The fossils date from about 80 million years ago to the end of the Cretaceous. However, the Apsaraviformes are likely older, because they seem to form an earlier branch than Gansus yumenensis, the earliest known member of Ornithurae. Gansus has been found in 120 million-year-old rocks in China.
One of the Ornithurines may have survived the end-Cretaceous extinction 66 million years ago. The bird in question is Qinornis paleocenica, found in the 61 million-year-old, Fangou formation in China (Xue, 1995). Although it has not be the subject of a phylogenetic analysis, Mayr (2007) argues that it is similar to Apsaravis.
The Hesperornithes (Fürbringer, 1888) were foot-propelled diving birds with teeth, sort of like loons with teeth, that were widespread in the northern hemisphere. The earliest fossils are about 100 million years old, and the latest come from the Lance formation at the end of the Cretaceous. Like the Enantiornithines, they were wiped out by the giant meteor impact at the end of the Cretaceous.
Carinatae Merrem, 1813
The Carinatae are the smallest clade containing Ichthyornis and modern birds. Ichthyornis was a toothed seabird of western North America, much of which was then covered by the Western Interior Seaway. The earliest known fossils date to about 95 million years ago. The latest relatives are known from the Lance formation at the end of the Cretaceous (Ornithurine D in Longrich et al., 2011).
Many of the birds from Wyoming's Lance formation seem to be very closely related to modern birds. The balance of the evidence is that none are modern birds (Longrich et al., 2011 and supplement), but they are very close.
One of the species studied by Longrich et al. (2011) is worthy of particular attention. The species hasn't been named yet, and is identified as Ornithurine C. It is a non-neorinthine carinatate bird, and it survived the Cretaceous extinction, at least for a time. Fossils of it are found in the Cretaceous Hell Creek (South Dakota and Montana) and Lance formations of Wyoming (prior to the extinction) and in the Paleocene Fort Union formation of Montana (after the extinction).
Neornithes Gadow, 1893
Finally we get to the modern birds. Neornithes consists of all birds descended from the most recent common ancestor of all extant bird species. Molecular evidence indicates that the division between Paleognaths and Neoganths is quite deep, much deeper than the division between either clade. For example, Mitchell et al. (2014) estimates the Paleoganth/Neognath split occurred 101 million years ago. The earliest split within the Paleognaths (Ostrich vs. the rest) is only 73 million years ago; the Galloanseres/Neoaves split is estimated at 89 million years ago, with none of the subsequent divisions more recent than 77 million years ago. Others get somewhat different numbers, but all agree that the Paleognath/Neognath division occurred long before any other.
If we take this at face value, one consequence is that for about 12 million years, there was only one lineage of modern bird, and that for another 12 million there were only 2. Further divisions did not occur until the last 11 million years of the Cretaceous. This paucity of modern birds explains how they can date back 100 million years ago without a trace in the fossil record. Indeed, most of the diversity is compressed into the last 6-7 million years of the Cretaceous. With such numbers, the most likely place to find Neornithe fossils is the Maastrichtian, and even in the Maastrichtian, you may still need to be lucky to find one. Birds near Neornithes are common enough, as Longrich et al. (2011) demonstrate, but actual Neornithes are scarce.
Other estimates based on Ericson et al. (2006a) or Brown et al. (2007) give better odds, but even the expansive estimate of Brown et al. (2008) only find 37 lineages surviving the extinction, although they suggest the number could be as high as 90. My own guess is that the lower numbers are closer to the truth as fossil support for higher numbers is quite weak.
It's interesting to note that Brown et al's point estimate for the Passeriform lineage is almost 90 million years ago. It was not long ago that ornithologists considered the passerines to be a recent lineage, and many reacted with disbelief when DNA data suggested they might be near the base of Neoaves. The tree presented here places the root of the Passeriform branch in a basal position in Passerae. The Passeriformes come at the end of our list because of our convention of putting the largest group last.
Cretaceous Fossil Neonithes
Although there are possible Cretaceous fossils of modern birds, most are pretty fragmentary and one can't be sure they belong to modern birds (Neornithes). There are plenty of fossils of other birds—Ichthyornithes, Hesperornithines, and Enantiornithes—just not Neornithes.
Should the lack of fossils be a big concern? No! The fossil record is incredibly sparse. How sparse? Caley (2007) gives a figure of around 2000 birds known only through fossils, compared with an estimate of 1.6 million bird species that have existed. That means we have only found fossils of 1 out of every 800 bird species.
How many Neornithes were there in the Cretaceous? They seem to have been a minor component of the avifauna, even at the end (Longrich et al., 2011) Let's try to get a ballpark number. Suppose Neonithes were 10% of the avian population and that overall diversity was similar to today—10,000 species. There would be 1000 Neornithes at the end of the Cretaceous. If we take an average species lifespan of 1 million years, and think of linear diversification over the previous 35 million years, we get a total of about 17,500 Mesozoic Neornithes. If we find 1 of every 8000, we would expect to find 2 fossil Mesozoic Neornithes. Needless to say, the estimate is pretty imprecise. However, it does indicate that true neorthine fossils will be rare.
It has also been suggested that location contributes to the problem. Since the vast majority of fossil collection has been in the northern hemisphere, one possible explanation for the paucity of supporting fossils is that most modern birds initially had a Gondwanan distribution (Cracraft, 2001; Ericson et al., 2002a). In that case, the lack of fossils further reflects low collection effort in the right location.
The best fossil candidate does come from Gondwana, more specifically, Antarctica. It is Vegavis iaai (Clarke et al., 2005). Clarke argues that Vegavis is very closely related to the Anatidae. It may even be in the Anatidae. Its existence implies that the Paleognaths, Galliformes, Anhimidae, and Anseranatidae were already separate lineages before the K/T extinction, as was at least one lineage in Neoaves.
Another possible Cretaceous neornithe is Austinornis lentus, from Texas. Clarke's (2004) phylogentic analysis placed it with the Galliformes, but evidence for this versus being a relatively basal member of Galloanseres is weak as it depends on a single charcter. Nonetheless, a position within Galloanseres was supported by a number of characters. Austinornis lived approximately 85 million years ago. According to Mitchell's (2014) analysis, this would be after the Galliformes/Neoaves split, but before the split between the Galliformes and the Anseriformes, which he places at about 73 million years ago.
Several other fossils that have been suggested as possible Neornithes fell just outside the Neornithes in Longrich et al. (2011). These include Ceramornis major, Cimolopteryx rara, "Cimolopteryx" maxima, "Cimolopteryx" minima, "Cimolopteryx" petra, and Iaceornis marshi. There are two at lease two other species in this group of near modern birds. Clarke (2004) found that Guildavis tener is there, while Slack et al. (2006) found it includes Apatornis celer. Most of the remaining possible fossil Neornithes are too fragmentary to even attempt a phylogentic analysis.
Diversification of Neornithes
Most of the Cretaceous neornithines were wiped out by the one-two whammy created by the Chicxulub meteor and Deccan traps volcanism. The situation for non-neornithine birds was even worse. Taking the error bars and misplaced passerines into account, Mitchell et al. (2014) estimate that a minimum of 4 neornithine lineages survived the giant meteor impact at the end of the Cretaceous: Paleognaths, Anseriformes, Galliformes, and Neoaves. It's possible that more than one neoavian survived.
My suspicion is that only four neoavian species survived, and that there was a rapid radiation once ecosystems started to recover in the Paleocene. This is consistent with the estimates by Ericsson et al. (2006a), which use a topology that is probably close to correct. I don't put much weight on Brown et al.'s (2008) estimates because they are based on a crazy topology. Similarly, Mitchell et al. (2014) have an inverted topology for Metaves/Coronaves, perhaps driven by the misplaced passerines, making those parts of their estimates unreliable.
The diversification of the surviving neornithes appears to have been rapid. Penguins had appeared by 62 million years ago, implying that the loon and seabird lineages had already split. There is uncertainty about whether some of the possibly more basal lineages—Opisthocomiformes, Cuculimorphae, and Musophagiformes—really are more basal. If they are, they must have split too. The earliest fossil owls are from the Paleocene (Burruornis). That implies at least 5 more separate lineages: Charadriiformes, Accipitrimorphae, mousebirds, Leptosomiformes-Piciformes, and Passerimorphae.
By the end of the Paleocene, Mitchell et al. (2014) estimate that all of the Paleognath orders had separated, except for Aepyornithiformes/Apterygiformes, which split in the early Eocene.
Schweizer et al. (2011) estimated the first split in the Psittaciformes during the mid-Paleocene. By then, the Cariamiformes, Falconiformes and Passeriformes must also have been independent lineages. The Passeriformes appear to be sister to the Zygodactalidae, which are known from early Eocene fossils. It is likely that the modern Passeriformes became a distinct lineage either in the Paleocene or possibly early Eocene. I consider attempts to anchor the modern Passeriformes at the beginning of the separation of New Zealand 82-85 million years ago to be dubious (e.g., Ericson et al., 2014).
By the end of the Eocene (about 34 million years ago), all of the modern bird orders (and many of the families) were distinct lineages. Of course, the Eocene avifauna also included many taxa that have no modern survivors, such as the Pelagornithidae (pseudotoothed birds), Sandcoleidae (sister to mousebirds), and Zygodactalidae (sister to Passeriformes).
Additional References
- Chiappe, L.M. (2007), “Glorified Dinosaurs: The Origin and Early Evolution of Birds”, Wiley, Hoboken, NJ.
- Chiappe, L.M., and G.J. Dyke (2002), The Mesozoic Radiation Of Birds, Annu. Rev. Ecol. Sys. 33, 91-124.
- Clarke, J.A. (2004), Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae), Bull. Amer. Mus. Nat. Hist. 286, 1-179.
- Clarke, J.A., C.P. Tambussi, J.I. Noriega, G.M. Erickson, and R.A. Ketcham (2005), Definitive fossil evidence for the extant avian radiation in the Cretaceous, Nature 433, 305-308.
- Ericson, P.G.P., S. Klopfstein, M. Irestedt, J.M.T. Nguyen, and J.A.A. Nylander (2014), Dating the diversification of the major lineages of Passeriformes (Aves), BMC Evo. Biol. 14, 8.
- Godefroit, P., A. Cau, H. Dong-Yu, F. Escuillié, W. Wenhao, and G. Dyke (2013), A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds, Nature 498, 359-362.
- Holtz, T.R. (2007), “Dinosaurs”, Random House, New York.
- Ksepka, D.T., and C.A. Boyd (2012), Quantifying historical trends in the completeness of the fossil record and the contributing factors: an example using Aves, Paleobiology 38, 112-125.
- Li, D., C. Sullivan, Z. Zhou, F. Zhang (2010), Basal birds from China: a brief review, Chinese Birds 1, 83-96.
- Longrich, N.R., T. Tokaryk, and D.J. Field (2011), Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary, Proc. Natl. Acad. Sci. 108, 15253-15257.
- Mayr, G. (2007), The Birds from the Paleocene Fissure Filling of Walbeck (Germany), J. Vert. Paleo. 27, 394-408.
- O'Connor, P.M., and C.A. Forster (2010), A Late Cretaceous (Maastrichtian) Avifauna from the Maevarano Formation, Madagascar, J. Vert. Paleo. 30, 1178-1201.
- Serrano, P.C. (1999), The Evolution of Dinosaurs, Science 284, 2137-2147.
- Slack, K.E., C.M. Jones,a T. Ando, G.L. Harrison, R.E. Fordyce, U. Arnason, and D. Penny (2006), Early Penguin Fossils, Plus Mitochondrial Genomes, Calibrate Avian Evolution, Mol. Biol. Evol. 23, 1144-1155.
- Xue, X. (1995), Qinornis paleocenica—A Paleocene bird discovered in China, Courier Forschungsinstitut Senckenberg 181, 89-93.
- You, H-L., M.C. Lamanna, J.D. Harris, L.M. Chiappe, J. O'Connor, S-A. Ji, J-C. Lu, C-X. Yuan, D-Q. Li, X. Zhang, K.J. Lacovara, P. Dodson, and Q. Ji (2006), A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China, Science 312, 1640-1643.
- Zhou, Z., J. Clarke, and F. Zhang (2008), Insight into diversity, body size and morphological evolution from the largest Early Cretaceous enantiornithine bird, J. Anat. 212, 565-577.
- Zhou, Z., and F.Z.Z. Li (2009), A new Lower Cretaceous bird from China and tooth reduction in early avian evolution, Proc. Royal Soc. B 277, 219-227.