APODIFORMES Peters, 1940 (1867)
This name has a complicated history. Priority would seem to indicate that Trochiliformes should be used as it dates back to Wagler, 1830 (as the suborder Trochili). However, modern usage is Apodiformes. For a long time, names based on Cypselus were used, but this is a junior subjective synonym of Apus, hence Apodiformes. For families, the updated name would retain the same priority as the original, which dates back to Huxley as Cypselomorphae. That's what the (1867) is about. Still, 1830 trumps 1867. In this case I think it's better to emphasize stability over priority and use Apodiformes, not Trochiliformes.
The Apodiformes are an old clade. Ksepka et al. (2013) have found a fossil Eocypselus from the Green River formation (about 48 milliion years old) from a lineage that seems to predate the swift/hummingbird split (the lineage, not necessarily the fossil).
Hemiprocnidae: Treeswifts Oberholser, 1906 (1852)
1 genus, 4 species HBW-5
- Crested Treeswift, Hemiprocne coronata
- Gray-rumped Treeswift, Hemiprocne longipennis
- Whiskered Treeswift, Hemiprocne comata
- Moustached Treeswift, Hemiprocne mystacea
Apodidae: Swifts Olphe-Galliard, 1887 (1836)
19 genera, 114 species HBW-5
The subfamilies and tribes follow HBW-5. Although they focus on the swiftlets, the available molecular studies suggest that some reorganization will be needed (see Thomassen et al., 2003, 2005; Price et al., 2004, 2005).
Based on Rheindt et al., the Glossy Swiftlet, Collocalia esculenta, has been split into 8 species:
- Tenggara Swiftlet, Collocalia sumbawae (sumbawae and sumbae)
- Christmas Island Swiftlet, Collocalia natalis (monotypic)
- Plume-toed Swiftlet, Collocalia affinis (affinis, elachyptera, vanderbilti, oberholseri, cyanoptila)
- Ridgetop Swiftlet, Collocalia isonota (isonota and bagobo)
- Grey-rumped Swiftlet, Collocalia marginata (septentrionalis and marginata)
- Drab Swiftlet, Collocalia neglecta (neglecta and perneglecta)
- Glossy Swiftlet, Collocalia esculenta (all other subspecies)
- Satin Swiftlet, Collocalia uropygialis (uropygialis and albidior)
- Spot-fronted Swift, Cypseloides cherriei
- White-chinned Swift, Cypseloides cryptus
- White-fronted Swift, Cypseloides storeri
- Black Swift / American Black Swift, Cypseloides niger
- White-chested Swift, Cypseloides lemosi
- Rothschild's Swift, Cypseloides rothschildi
- Sooty Swift, Cypseloides fumigatus
- Great Dusky Swift, Cypseloides senex
- Chestnut-collared Swift, Streptoprocne rutila
- Tepui Swift, Streptoprocne phelpsi
- White-collared Swift, Streptoprocne zonaris
- Biscutate Swift, Streptoprocne biscutata
- White-naped Swift, Streptoprocne semicollaris
- Pygmy Swiftlet, Collocalia troglodytes
- Tenggara Swiftlet, Collocalia sumbawae
- Bornean Swiftlet, Collocalia dodgei
- Cave Swiftlet, Collocalia linchi
- Christmas Island Swiftlet, Collocalia natalis
- Plume-toed Swiftlet, Collocalia affinis
- Ridgetop Swiftlet, Collocalia isonota
- Grey-rumped Swiftlet, Collocalia marginata
- Drab Swiftlet, Collocalia neglecta
- Glossy Swiftlet, Collocalia esculenta
- Satin Swiftlet, Collocalia uropygialis
- Giant Swiftlet, Hydrochous gigas
- Three-toed Swiftlet, Hydrochous papuensis
- Black-nest Swiftlet, Aerodramus maximus
- Indian Swiftlet, Aerodramus unicolor
- Himalayan Swiftlet, Aerodramus brevirostris
- Volcano Swiftlet, Aerodramus vulcanorum
- Whitehead's Swiftlet, Aerodramus whiteheadi
- Bare-legged Swiftlet, Aerodramus nuditarsus
- Mayr's Swiftlet, Aerodramus orientalis
- Halmahera Swiftlet, Aerodramus infuscatus
- Sulawesi Swiftlet, Aerodramus sororum
- Seram Swiftlet, Aerodramus ceramensis
- Mountain Swiftlet, Aerodramus hirundinaceus
- Australian Swiftlet, Aerodramus terraereginae
- Uniform Swiftlet, Aerodramus vanikorensis
- Marquesan Swiftlet, Aerodramus ocistus
- Tahiti Swiftlet, Aerodramus leucophaeus
- Atiu Swiftlet, Aerodramus sawtelli
- Island Swiftlet, Aerodramus inquietus
- Palau Swiftlet, Aerodramus pelewensis
- Mariana Swiftlet, Aerodramus bartschi
- Ameline Swiftlet, Aerodramus amelis
- Philippine Swiftlet, Aerodramus mearnsi
- White-rumped Swiftlet, Aerodramus spodiopygius
- Seychelles Swiftlet, Aerodramus elaphrus
- Mascarene Swiftlet, Aerodramus francicus
- Germain's Swiftlet, Aerodramus germani
- Edible-nest Swiftlet, Aerodramus fuciphagus
- Mossy-nest Swiftlet, Aerodramus salangana
- Scarce Swift, Schoutedenapus myoptilus
- Schouteden's Swift, Schoutedenapus schoutedeni
- Philippine Spinetailed-Swift, Mearnsia picina
- Papuan Spinetailed-Swift, Mearnsia novaeguineae
- Madagascan Spinetail, Zoonavena grandidieri
- Sao Tome Spinetail, Zoonavena thomensis
- White-rumped Spinetail, Zoonavena sylvatica
- Mottled Spinetail, Telacanthura ussheri
- Black Spinetail, Telacanthura melanopygia
- Silver-rumped Spinetail, Rhaphidura leucopygialis
- Sabine's Spinetail, Rhaphidura sabini
- Cassin's Spinetail, Neafrapus cassini
- Boehm's Spinetail, Neafrapus boehmi
- White-throated Needletail, Hirundapus caudacutus
- Silver-backed Needletail, Hirundapus cochinchinensis
- Brown-backed Needletail, Hirundapus giganteus
- Purple Needletail, Hirundapus celebensis
- Lesser Antillean Swift, Chaetura martinica
- Band-rumped Swift, Chaetura spinicaudus
- Costa Rican Swift, Chaetura fumosa
- Gray-rumped Swift, Chaetura cinereiventris
- Pale-rumped Swift, Chaetura egregia
- Vaux's Swift, Chaetura vauxi
- Chimney Swift, Chaetura pelagica
- Chapman's Swift, Chaetura chapmani
- Amazonian Swift / Mato Grosso Swift, Chaetura viridipennis
- Sick's Swift, Chaetura meridionalis
- Short-tailed Swift, Chaetura brachyura
- Tumbes Swift, Chaetura ocypetes
- White-throated Swift, Aeronautes saxatalis
- White-tipped Swift, Aeronautes montivagus
- Andean Swift, Aeronautes andecolus
- Antillean Palm-Swift, Tachornis phoenicobia
- Pygmy Swift / Pygmy Palm-Swift, Tachornis furcata
- Fork-tailed Palm-Swift / Neotropical Palm-Swift, Tachornis squamata
- Lesser Swallow-tailed Swift, Panyptila cayennensis
- Great Swallow-tailed Swift, Panyptila sanctihieronymi
- African Palm-Swift, Cypsiurus parvus
- Asian Palm-Swift, Cypsiurus balasiensis
- Alpine Swift, Tachymarptis melba
- Mottled Swift, Tachymarptis aequatorialis
- Dark-rumped Swift, Apus acuticauda
- Cook's Swift, Apus cooki
- Blyth's Swift, Apus leuconyx
- Salim Ali's Swift, Apus salimalii
- Pacific Swift, Apus pacificus
- White-rumped Swift, Apus caffer
- Bates's Swift, Apus batesi
- Horus Swift, Apus horus
- Little Swift, Apus affinis
- House Swift, Apus nipalensis
- Nyanza Swift, Apus niansae
- Malagasy Black-Swift, Apus balstoni
- Bradfield's Swift, Apus bradfieldi
- African Black-Swift, Apus barbatus
- Fernando Po Swift, Apus sladeniae
- Cape Verde Swift, Apus alexandri
- Forbes-Watson's Swift, Apus berliozi
- Plain Swift, Apus unicolor
- Common Swift, Apus apus
- Pallid Swift, Apus pallidus
- Eulampis (Boie 1831) has been merged into Anthracothorax (Boie 1831). Priority was determined by the first reviser action of Remsen et al. (2015).
- Chalcostigma (Reichenbach 1854) and Oreonympha (Gould 1869) have been merged into Oxypogon (Gould 1848).
- The Marvelous Spatuletail (Loddigesia Bonaparte 1850) has been merged into Eriocnemis (Reichenbach 1849). This causes a name conflict as two species are mirabilis. The Marvelous Spatuletail gets to keep the name, and the Colorful Puffleg needs a new name. As none is available, I refer to it as Eriocnemis "mirabilis" for now.
- Clytolaema (Gould 1853) has been merged into Heliodoxa (Gould 1850).
- Nesophlox (Ridgway 1910) and Philodice (Mulsant, Verreaux and Verreaux 1866, type mitchellii) have been separated from Calliphlox, which is left with a single species. Note that Nesophlox may consist of two species (Feo et al., 2015).
- Atthis (Reichenbach 1854) has been merged into Selasphorus (Swainson 1832). Selasphorus already includes Stellula (AOU supplement 53 and McGuire et al., 2009).
- Basilinna (Boie 1831, type leucotis) has been separated from Hylocharis.
- Pampa (Reichenbach 1854, type pampa) has been separated from Campylopterus.
- Chlorostilbon (Gould 1853) and Cyanophaia (Reichenbach 1854) have been merged into Cynanthus (Swainson 1827).
- The Mexican Woodnymph has been moved to Eupherusa from Thalurania..
- Talaphorus (Simon 1910, type hypostictus) and Thaumasius (Sclater 1879, type taczanowskii) have been separated from Leucippus, which is now reduced to a single species.
- Saucerottia (Bonaparte 1850, type saucerrottei) has been separated from Amazilia.
- Lepidopyga (Reichenbach 1855) has been merged into Chrysuronia (Bonaparte 1850), which has been separated from Amazilia and has also gained two former members of Hylocharis.
- The remaining Hylocharis have been augmented with some former members of Amazilia.
- Polyerata (Heine 1863, type amabilis) has been separated from Amazilia.
- Chlorestes (Reichenbach 1854, type notata) has been augmented with Juliamyia (not Damophila Reichenbach 1854, which is preoccupied by the moth genus Damophila Curtis 1832), two former Hylocharis and one former Amazilia.
- Fiery Topaz, Topaza pyra
Click for Topazes & Hermits tree - Crimson Topaz, Topaza pella
- White-necked Jacobin, Florisuga mellivora
- Black Jacobin, Florisuga fusca
- White-tipped Sicklebill, Eutoxeres aquila
- Buff-tailed Sicklebill, Eutoxeres condamini
- Saw-billed Hermit, Ramphodon naevius
- Bronzy Hermit, Glaucis aeneus
- Rufous-breasted Hermit, Glaucis hirsutus
- Hook-billed Hermit, Glaucis dohrnii
- Band-tailed Barbthroat, Threnetes ruckeri
- Pale-tailed Barbthroat, Threnetes leucurus
- Sooty Barbthroat, Threnetes niger
- Broad-tipped Hermit, Anopetia gounellei
- White-bearded Hermit, Phaethornis hispidus
- Tawny-bellied Hermit, Phaethornis syrmatophorus
- Mexican Hermit, Phaethornis mexicanus
- Long-billed Hermit, Phaethornis longirostris
- Green Hermit, Phaethornis guy
- White-whiskered Hermit, Phaethornis yaruqui
- Great-billed Hermit, Phaethornis malaris
- Long-tailed Hermit, Phaethornis superciliosus
- Straight-billed Hermit, Phaethornis bourcieri
- Koepcke's Hermit, Phaethornis koepckeae
- Needle-billed Hermit, Phaethornis philippii
- Buff-bellied Hermit, Phaethornis subochraceus
- Scale-throated Hermit, Phaethornis eurynome
- Sooty-capped Hermit, Phaethornis augusti
- Planalto Hermit, Phaethornis pretrei
- Pale-bellied Hermit, Phaethornis anthophilus
- Stripe-throated Hermit, Phaethornis striigularis
- Gray-chinned Hermit, Phaethornis griseogularis
- Black-throated Hermit, Phaethornis atrimentalis
- Reddish Hermit, Phaethornis ruber
- White-browed Hermit, Phaethornis stuarti
- Dusky-throated Hermit, Phaethornis squalidus
- Streak-throated Hermit, Phaethornis rupurumii
- Cinnamon-throated Hermit, Phaethornis nattereri
- Little Hermit, Phaethornis longuemareus
- Tapajos Hermit, Phaethornis aethopygus
- Minute Hermit, Phaethornis idaliae
- Green-fronted Lancebill, Doryfera ludovicae
Click for Mango tree - Blue-fronted Lancebill, Doryfera johannae
- Wedge-billed Hummingbird, Schistes geoffroyi
- Hooded Visorbearer, Augastes lumachella
- Hyacinth Visorbearer, Augastes scutatus
- White-vented Violetear, Colibri serrirostris
- Sparkling Violetear, Colibri coruscans
- Brown Violetear, Colibri delphinae
- Mexican Violetear, Colibri thalassinus
- Lesser Violetear, Colibri cyanotus
- Horned Sungem, Heliactin bilophus
- Tooth-billed Hummingbird, Androdon aequatorialis
- Purple-crowned Fairy, Heliothryx barroti
- Black-eared Fairy, Heliothryx auritus
- White-tailed Goldenthroat, Polytmus guainumbi
- Green-tailed Goldenthroat, Polytmus theresiae
- Tepui Goldenthroat, Polytmus milleri
- Fiery-tailed Awlbill, Avocettula recurvirostris
- Ruby-topaz Hummingbird, Chrysolampis mosquitus
- Jamaican Mango, Anthracothorax mango
- Black-throated Mango, Anthracothorax nigricollis
- Green-throated Mango, Anthracothorax viridigula
- Green-breasted Mango, Anthracothorax prevostii
- Veraguan Mango, Anthracothorax veraguensis
- Antillean Mango, Anthracothorax dominicus
- Green Mango, Anthracothorax viridis
- Green-throated Carib, Anthracothorax holosericeus
- Purple-throated Carib, Anthracothorax jugularis
- Green-backed Firecrown, Sephanoides sephaniodes
Click for Coquette tree - Juan Fernandez Firecrown, Sephanoides fernandensis
- Racket-tailed Coquette, Discosura longicaudus
- Green Thorntail, Discosura conversii
- Wire-crested Thorntail, Discosura popelairii
- Black-bellied Thorntail, Discosura langsdorffi
- Coppery Thorntail / Letitia's Thorntail, Discosura letitiae
- Tufted Coquette, Lophornis ornatus
- Dot-eared Coquette, Lophornis gouldii
- Frilled Coquette, Lophornis magnificus
- Black-crested Coquette, Lophornis helenae
- White-crested Coquette, Lophornis adorabilis
- Peacock Coquette, Lophornis pavoninus
- Festive Coquette, Lophornis chalybeus
- Short-crested Coquette, Lophornis brachylophus
- Rufous-crested Coquette, Lophornis delattrei
- Spangled Coquette, Lophornis stictolophus
- Ecuadorian Piedtail, Phlogophilus hemileucurus
- Peruvian Piedtail, Phlogophilus harterti
- Royal Sunangel, Heliangelus regalis
- Gorgeted Sunangel, Heliangelus strophianus
- Orange-throated Sunangel, Heliangelus mavors
- Amethyst-throated Sunangel, Heliangelus amethysticollis
- Little Sunangel / Flame-throated Sunangel, Heliangelus micraster
- Tourmaline Sunangel, Heliangelus exortis
- Purple-throated Sunangel, Heliangelus viola
- Speckled Hummingbird, Adelomyia melanogenys
- Gray-bellied Comet, Taphrolesbia griseiventris
- Long-tailed Sylph, Aglaiocercus kingii
- Violet-tailed Sylph, Aglaiocercus coelestis
- Venezuelan Sylph, Aglaiocercus berlepschi
- Red-tailed Comet, Sappho sparganurus
- Black-tailed Trainbearer, Lesbia victoriae
- Green-tailed Trainbearer, Lesbia nuna
- Black-backed Thornbill, Ramphomicron dorsale
- Purple-backed Thornbill, Ramphomicron microrhynchum
- Wedge-tailed Hillstar, Oreotrochilus adela
- Andean Hillstar, Oreotrochilus estella
- Ecuadorian Hillstar, Oreotrochilus chimborazo
- Black-breasted Hillstar, Oreotrochilus melanogaster
- White-sided Hillstar, Oreotrochilus leucopleurus
- Mountain Avocetbill, Opisthoprora euryptera
- Bronze-tailed Comet, Polyonymus caroli
- Rainbow-bearded Thornbill, Oxypogon herrani
- Rufous-capped Thornbill, Oxypogon ruficeps
- Olivaceous Thornbill, Oxypogon olivaceus
- Bearded Mountaineer, Oxypogon nobilis
- Bronze-tailed Thornbill, Oxypogon heteropogon
- Blue-mantled Thornbill, Oxypogon stanleyi
- Blue-bearded Helmetcrest, Oxypogon cyanolaemus
- White-bearded Helmetcrest, Oxypogon lindenii
- Buffy Helmetcrest, Oxypogon stuebelii
- Green-bearded Helmetcrest, Oxypogon guerinii
- Black Metaltail, Metallura phoebe
- Tyrian Metaltail, Metallura tyrianthina
- Perija Metaltail, Metallura iracunda
- Scaled Metaltail, Metallura aeneocauda
- Coppery Metaltail, Metallura theresiae
- Fire-throated Metaltail / Fiery-throated Metaltail, Metallura eupogon
- Neblina Metaltail, Metallura odomae
- Viridian Metaltail, Metallura williami
- Violet-throated Metaltail, Metallura baroni
- Hoary Puffleg, Haplophaedia lugens
Click for Brilliants tree - Greenish Puffleg, Haplophaedia aureliae
- Buff-thighed Puffleg, Haplophaedia assimilis
- Colorful Puffleg, Eriocnemis "mirabilis"
- Emerald-bellied Puffleg, Eriocnemis aline
- Sapphire-vented Puffleg, Eriocnemis luciani
- Black-thighed Puffleg, Eriocnemis derbyi
- Coppery-bellied Puffleg, Eriocnemis cupreoventris
- Turquoise-throated Puffleg, Eriocnemis godini
- Marvelous Spatuletail, Eriocnemis mirabilis
- Blue-capped Puffleg, Eriocnemis glaucopoides
- Golden-breasted Puffleg, Eriocnemis mosquera
- Glowing Puffleg, Eriocnemis vestita
- Gorgeted Puffleg, Eriocnemis isabellae
- Black-breasted Puffleg, Eriocnemis nigrivestis
- Mountain Velvetbreast, Lafresnaya lafresnayi
- Shining Sunbeam, Aglaeactis cupripennis
- Purple-backed Sunbeam, Aglaeactis aliciae
- White-tufted Sunbeam, Aglaeactis castelnaudii
- Black-hooded Sunbeam, Aglaeactis pamela
- Black Inca, Coeligena prunellei
- Bronzy Inca, Coeligena coeligena
- Brown Inca, Coeligena wilsoni
- Collared Inca, Coeligena torquata
- Violet-throated Starfrontlet, Coeligena violifer
- Rainbow Starfrontlet, Coeligena iris
- White-tailed Starfrontlet, Coeligena phalerata
- Dusky Starfrontlet, Coeligena orina
- Buff-winged Starfrontlet, Coeligena lutetiae
- Golden-bellied Starfrontlet, Coeligena bonapartei
- Blue-throated Starfrontlet, Coeligena helianthea
- Sword-billed Hummingbird, Ensifera ensifera
- Great Sapphirewing, Pterophanes cyanopterus
- Velvet-purple Coronet, Boissonneaua jardini
- Buff-tailed Coronet, Boissonneaua flavescens
- Chestnut-breasted Coronet, Boissonneaua matthewsii
- Booted Racket-tail, Ocreatus underwoodii
- Purple-bibbed Whitetip, Urosticte benjamini
- Rufous-vented Whitetip, Urosticte ruficrissa
- White-tailed Hillstar, Urochroa bougueri
- Violet-chested Hummingbird, Sternoclyta cyanopectus
- Scissor-tailed Hummingbird, Hylonympha macrocerca
- Black-throated Brilliant, Heliodoxa schreibersii
- Velvet-browed Brilliant, Heliodoxa xanthogonys
- Pink-throated Brilliant, Heliodoxa gularis
- Rufous-webbed Brilliant, Heliodoxa branickii
- Gould's Jewelfront, Heliodoxa aurescens
- Brazilian Ruby, Heliodoxa rubricauda
- Green-crowned Brilliant, Heliodoxa jacula
- Violet-fronted Brilliant, Heliodoxa leadbeateri
- Fawn-breasted Brilliant, Heliodoxa rubinoides
- Empress Brilliant, Heliodoxa imperatrix
- Giant Hummingbird, Patagona gigas
Click for Giant Hummingbird,
Mountain-gems,
and Bees tree - Garnet-throated Hummingbird, Lamprolaima rhami
- Magnificent Hummingbird / Rivoli's Hummingbird, Eugenes fulgens
- Admirable Hummingbird, Eugenes spectabilis
- Fiery-throated Hummingbird, Panterpe insignis
- Plain-capped Starthroat, Heliomaster constantii
- Long-billed Starthroat, Heliomaster longirostris
- Stripe-breasted Starthroat, Heliomaster squamosus
- Blue-tufted Starthroat, Heliomaster furcifer
- White-bellied Mountain-gem, Lampornis hemileucus
- Blue-throated Hummingbird / Blue-throated Mountain-gem, Lampornis clemenciae
- Amethyst-throated Hummingbird / Amethyst-throated Mountain-gem, Lampornis amethystinus
- Green-throated Mountain-gem, Lampornis viridipallens
- Green-breasted Mountain-gem, Lampornis sybillae
- Gray-tailed Mountain-gem, Lampornis cinereicauda
- Purple-throated Mountain-gem, Lampornis calolaemus
- White-throated Mountain-gem, Lampornis castaneoventris
- Sparkling-tailed Hummingbird / Sparkling-tailed Woodstar, Tilmatura dupontii
- Amethyst Woodstar, Calliphlox amethystina
- Oasis Hummingbird, Rhodopis vesper
- Purple-collared Woodstar, Myrtis fanny
- Short-tailed Woodstar, Myrmia micrura
- Peruvian Sheartail, Thaumastura cora
- Chilean Woodstar, Eulidia yarrellii
- Magenta-throated Woodstar, Philodice bryantae
- Purple-throated Woodstar, Philodice mitchellii
- Slender-tailed Woodstar, Microstilbon burmeisteri
- Rufous-shafted Woodstar, Chaetocercus jourdanii
- White-bellied Woodstar, Chaetocercus mulsant
- Little Woodstar, Chaetocercus bombus
- Gorgeted Woodstar, Chaetocercus heliodor
- Santa Marta Woodstar, Chaetocercus astreans
- Esmeraldas Woodstar, Chaetocercus berlepschi
- Mexican Sheartail, Doricha eliza
- Slender Sheartail, Doricha enicura
- Lucifer Hummingbird / Lucifer Sheartail, Calothorax lucifer
- Beautiful Hummingbird / Beautiful Sheartail, Calothorax pulcher
- Black-chinned Hummingbird, Archilochus alexandri
- Ruby-throated Hummingbird, Archilochus colubris
- Bahama Woodstar, Nesophlox evelynae
- Inagua Woodstar, Nesophlox lyrura
- Bee Hummingbird, Mellisuga helenae
- Vervain Hummingbird, Mellisuga minima
- Anna's Hummingbird, Calypte anna
- Costa's Hummingbird, Calypte costae
- Broad-tailed Hummingbird, Selasphorus platycercus
- Rufous Hummingbird, Selasphorus rufus
- Calliope Hummingbird, Selasphorus calliope
- Allen's Hummingbird, Selasphorus sasin
- Bumblebee Hummingbird, Selasphorus heloisa
- Wine-throated Hummingbird, Selasphorus ellioti
- Volcano Hummingbird, Selasphorus flammula
- Scintillant Hummingbird, Selasphorus scintilla
- Glow-throated Hummingbird, Selasphorus ardens
- White-eared Hummingbird, Basilinna leucotis
Click for Emeralds tree - Xantus's Hummingbird, Basilinna xantusii
- Wedge-tailed Sabrewing, Pampa pampa
- Curve-winged Sabrewing, Pampa curvipennis
- Long-tailed Sabrewing, Pampa excellens
- Emerald-chinned Hummingbird, Abeillia abeillei
- Violet-headed Hummingbird, Klais guimeti
- Antillean Crested Hummingbird, Orthorhyncus cristatus
- Santa Marta Blossomcrown, Anthocephala floriceps
- Tolima Blossomcrown, Anthocephala berlepschi
- Purple-crowned Plovercrest, Stephanoxis loddigesii
- Green-crowned Plovercrest, Stephanoxis lalandi
- Gray-breasted Sabrewing, Campylopterus largipennis
- Dry Forest Sabrewing, Campylopterus calcirupicola
- Violet Sabrewing, Campylopterus hemileucurus
- Rufous Sabrewing, Campylopterus rufus
- Buff-breasted Sabrewing, Campylopterus duidae
- Rufous-breasted Sabrewing, Campylopterus hyperythrus
- Napo Sabrewing, Campylopterus villaviscensio
- Santa Marta Sabrewing, Campylopterus phainopeplus
- Lazuline Sabrewing, Campylopterus falcatus
- White-tailed Sabrewing, Campylopterus ensipennis
- Dusky Hummingbird, Cynanthus sordidus
- Blue-headed Hummingbird, Cynanthus bicolor
- Brace's Emerald, Cynanthus bracei
- Cuban Emerald, Cynanthus ricordii
- Caribbean Emerald / Gould's Emerald, Cynanthus elegans
- Hispaniolan Emerald, Cynanthus swainsonii
- Puerto Rican Emerald, Cynanthus maugaeus
- Coppery Emerald, Cynanthus russatus
- Broad-billed Hummingbird, Cynanthus latirostris
- Doubleday's Hummingbird, Cynanthus doubledayi
- Golden-crowned Emerald, Cynanthus auriceps
- Cozumel Emerald, Cynanthus forficatus
- Canivet's Emerald, Cynanthus canivetii
- Garden Emerald, Cynanthus assimilis
- Western Emerald, Cynanthus melanorhynchus
- Green-tailed Emerald, Cynanthus alice
- Narrow-tailed Emerald, Cynanthus stenurus
- Short-tailed Emerald, Cynanthus poortmani
- Glittering-bellied Emerald, Cynanthus lucidus
- Blue-tailed Emerald, Cynanthus mellisugus
- Red-billed Emerald, Cynanthus gibsoni
- Chiribiquete Emerald, Cynanthus olivaresi
- Bronze-tailed Plumeleteer, Chalybura urochrysia
Click for Amazilias tree - White-vented Plumeleteer, Chalybura buffonii
- Violet-capped Woodnymph, Thalurania glaucopis
- Long-tailed Woodnymph, Thalurania watertonii
- Crowned Woodnymph, Thalurania colombica
- Fork-tailed Woodnymph, Thalurania furcata
- Snowcap, Microchera albocoronata
- Coppery-headed Emerald, Elvira cupreiceps
- White-tailed Emerald, Elvira chionura
- Violet-capped Hummingbird, Goldmania violiceps
- Pirre Hummingbird, Goethalsia bella
- Mexican Woodnymph, Eupherusa ridgwayi
- White-tailed Hummingbird, Eupherusa poliocerca
- Blue-capped Hummingbird / Oaxaca Hummingbird, Eupherusa cyanophrys
- Stripe-tailed Hummingbird, Eupherusa eximia
- Black-bellied Hummingbird, Eupherusa nigriventris
- Scaly-breasted Hummingbird, Phaeochroa cuvierii
- Buffy Hummingbird, Leucippus fallax
- Tumbes Hummingbird, Thaumasius baeri
- Spot-throated Hummingbird, Thaumasius taczanowskii
- Many-spotted Hummingbird, Taphrospilus hypostictus
- Swallow-tailed Hummingbird, Eupetomena macroura
- Sombre Hummingbird, Aphantochroa cirrochloris
- Olive-spotted Hummingbird, Talaphorus chlorocercus
- Red-billed Streamertail / Streamertail, Trochilus polytmus
- Black-billed Streamertail, Trochilus scitulus
- Violet-crowned Hummingbird, Saucerottia violiceps
- Green-fronted Hummingbird, Saucerottia viridifrons
- Azure-crowned Hummingbird, Saucerottia cyanocephala
- Blue-vented Hummingbird, Saucerottia hoffmanni
- Berylline Hummingbird, Saucerottia beryllina
- Blue-tailed Hummingbird, Saucerottia cyanura
- Snowy-bellied Hummingbird, Saucerottia edward
- Steely-vented Hummingbird, Saucerottia saucerottei
- Indigo-capped Hummingbird, Saucerottia cyanifrons
- Chestnut-bellied Hummingbird, Saucerottia castaneiventris
- Green-bellied Hummingbird, Saucerottia viridigaster
- Copper-rumped Hummingbird, Saucerottia tobaci
- Cinnamon Hummingbird, Amazilia rutila
- Buff-bellied Hummingbird, Amazilia yucatanensis
- Rufous-tailed Hummingbird, Amazilia tzacatl
- Amazilia Hummingbird, Chrysuronia amazilia
- Andean Emerald, Chrysuronia franciae
- Versicolored Emerald, Chrysuronia versicolor
- Shining-green Hummingbird, Chrysuronia goudoti
- Golden-tailed Sapphire, Chrysuronia oenone
- Sapphire-throated Hummingbird, Chrysuronia coeruleogularis
- Sapphire-bellied Hummingbird, Chrysuronia lilliae
- Humboldt's Sapphire, Chrysuronia humboldtii
- Blue-headed Sapphire, Chrysuronia grayi
- White-chested Emerald, Chrysuronia brevirostris
- Plain-bellied Emerald, Chrysuronia leucogaster
- White-throated Hummingbird, Leucochloris albicollis
- Glittering-throated Emerald, Hylocharis fimbriata
- Sapphire-spangled Emerald, Hylocharis lactea
- Rufous-throated Sapphire, Hylocharis sapphirina
- Gilded Hummingbird / Gilded Sapphire, Hylocharis chrysura
- White-bellied Hummingbird, Hylocharis chionogaster
- Green-and-white Hummingbird, Hylocharis viridicauda
- Honduran Emerald, Polyerata luciae
- Mangrove Hummingbird, Polyerata boucardi
- Purple-chested Hummingbird, Polyerata rosenbergi
- Blue-chested Hummingbird, Polyerata amabilis
- Charming Hummingbird, Polyerata decora
- White-bellied Emerald, Chlorestes candida
- Blue-throated Goldentail / Blue-throated Sapphire, Chlorestes eliciae
- White-chinned Sapphire, Chlorestes cyanus
- Violet-bellied Hummingbird, Chlorestes julie
- Blue-chinned Sapphire, Chlorestes notata
The Ameline Swiftlet, Aerodramus amelis, including palawanensis, has been split from the Uniform Swiftlet, Aerodramus vanikorensis (see Price et al., 2005; Dickinson and Remsen, 2013). The Three-toed Swiftlet seems to be sister to the Giant Swiftlet, Hydrochous gigas (Price et al., 2005), and so has been moved to Hydrochous.
Tumbes Swift, Chaetura ocypetes, has been split from Short-tailed Swift, Chaetura brachyura, as in Ridgely and Greenfield (2001).
Based on Päckert et al. (2012a), Alpine Swift, Tachymarptis melba and Mottled Swift, Tachymarptis aequatorialis, have been returned to Tachymarptis (from Apus). Further, Apus has been rearranged based on their study.
Following Leader (2011), the Fork-tailed Swift, Apus pacificus, has been split into 4 species: Blyth's Swift, Apus leuconyx, Salim Ali's Swift, Apus salimalii, Pacific Swift, Apus pacificus, and Cook's Swift, Apus cooki. In fact, Päckert et al. (2012a) subsequently found that Cook's Swift is more closely related to the Dark-rumped Swift than to the Pacific Swift. They did not include Blyth's or Salim Ali's Swifts, but it is likely that they are closer to Pacific, and that the Dark-rumped/Pacific complexes are themselves sisters.
Cypseloidinae: Primitive American Swifts Brooke, 1970
Apodinae Olphe-Galliard, 1887 (1836)
Collocalini: Swiftlets Bonaparte, 1853 (1852)
Chaeturini: Needletails Bonaparte, 1857
Apodini: Typical Swifts Olphe-Galliard, 1887 (1836)
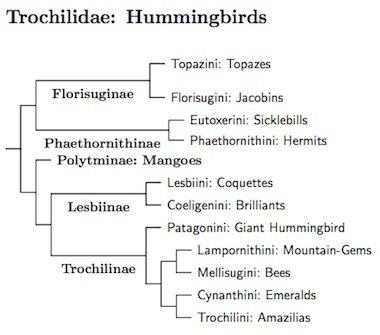
Trochilidae: Hummingbirds Vigors, 1825
103 genera, 350 species HBW-5
 |
| Click for Trochilidae genera |
|---|
Hummingbird taxonomy has been substantially revised in the 21st century. Past editions of this page have been based on Altshuler et al. (2004), McGuire et al. (2007, 2009), and Kirchman et al. (2010). It is now based on the phylogeny of McGuire et al. (2014) which includes 85% of all hummingbird species. Although some of the details differ, the AOU's SACC has also adopted this type of arrangement.
The subfamilies and tribes represent some of the natural groupings in McGuire et al. (2014). The treatment here is similar to that of H&M-4 (Dickinson and Remsen, 2013), but differs in several respects. I have acknowledged the deep divisions within Florisuginae and Phaethornithinae by dividing them into two tribes each, Topazini (Topazes) and Florisugini (Jacobins), and Eutoxerini (Sicklebills) and Phaethornithini (Hermits). I treat the Giant Hummingbird as a monotypic tribe (Patagonini) within Trochilinae rather than a subfamily sister to Trochilinae. And last, I've separated the core emeralds (Cynanthini) from the Amazilia group (Trochilini). The time-calibrated tree in McGuire et al. (2014) suggests that all three subfamilies originated about 20-22 mya and that the tribes from 12-20 mya. I suspect that the timing on their tree is a somewhat compressed, even though the swift-hummingbird split agrees with Jarvis et al. (2014). Ksepka and Clarke's (2015) analysis of the fossil record concludes that the swift-hummingbird split occurred earlier, no later than 51 mya.
There are also fairly deep divisions in the mangos, between the lancebill-violetear clade, the sungem-fairy clade, and the rest. These could be recognized as tribes—Petasophorini (violetears, based on Petasophora = Colibri), Heliothrichini(fairies), and Polytmini (mangos). However, I'm not sure that doing so really adds that much information.
There is an issue concerning one of of the tribe names. I've used Coeligenini (Eudes-Deslongchamps 1881) and as if it were a pre-1961 replacement name for Heliantheini (Reichenbach 1853). The problem is that until 2009, neither name was in prevailing usage, so that article 40.2 of the ICZN Code does not really apply. The problem would be even worse if Docimastini (Reichenbach 1853 based on Docimastes Gould 1849 = Ensifera Lesson 1843) had not been eliminated from consideration by the first reviser action of Dickinson and Remsen (2013).
McGuire et al. (2009) proposed use of Coeligenini as a Phylocode clade name. This has gained some traction. To avoid confusion, I also use Coeligenini and treat it a replacement for the name Heliantheini, although it may not stricly follow the the Code. If that bothers you, think of it as a Phylocode clade name (other such names are used elsewhere). Since Dickinson and Remsen (2013) adopted Heliantheini, it may come back into common use, in which case I will reconsider.
Hummingbird Generic Changes
Compared to H&M-4 (Dickinson and Remsen, 2013), there have been a number of genus changes in the hummingbirds:
Hummingbird Hybrid
Pérez-Emán et al. (2017) found that the Bogota Sunangel was actually a hybrid. Kirchman et al.'s (2010) finding that it was related to Sylphs was incorrect due to considering insufficient mitochondrial DNA from sylph species. Broader sampling revealed that the Bogota Sunangel shares a haplotype with the Long-tailed Sylph, Aglaiocercus kingii. This is a strong indication that its mother was a Long-tailed Sylph, and that the Bogota Sunangel was a hybrid.
Hummingbird Splits and Lumps
The Mexican Hermit, Phaethornis mexicanus, has been spilt from Long-billed Hermit, Phaethornis longirostris. See Howell (2013), and Arbeláz-Cortés and Navarro-Sigüenza (2013).
The Green Violetear, Colibri thalassinus, has been split into Mexican Violetear, Colibri thalassinus, and Lesser Violetear, Colibri cyanotus based on Remsen et al. (2015).
The Speckled Hummingbird, Adelomyia melanogenys, may include 5-6 species, one of them unnamed (Chaves and Smith, 2011; Chaves et al., 2011).
The Bearded Helmetcrest, Oxypogon guerinii has been split into: Green-bearded Helmetcrest, Oxypogon guerinii, Blue-bearded Helmetcrest, Oxypogon cyanolaemus, White-bearded Helmetcrest, Oxypogon lindenii, and Buffy Helmetcrest, Oxypogon stuebelii based on Collar and Salaman (2013).
It has long been argued that the distinctive Admirable Hummingbird, Eugenes spectabilis, of Costa Rica and Panama should be considered a separate species from the more northern Magnificent Hummingbird, Eugenes fulgens. Indeed, some authors have split them. Now there is genetic evidence to support this. Zamudio-Beltran et al. (2015) Admirable separated by a healthy 6 percent mitochondrial genetic distance. They also suggest subdividing fulgens and giving viridiceps species status. I don't see that they really provide evidence for this. They find that fulgens and viridiceps are not reciprocally monophyletic (strike 1), that there are no known plumage distinctions (strike 2), and that the mitochondrial genetic distance between fulgens and the viridiceps is relatively modest, less that 1 percent (strike 3).
 |
| Purple-throated Mountain-gem Lampornis calolaemus |
|---|
| Monteverde, Costa Rica, 2003 |
The proper taxonomic treatment of the Lampornis castaneoventris complex remains unclear. There are three forms: with purple throats and blue tails (calolaemus, pectoralis, and homogenes), with white throats and blue tails (castaneoventris), and with white throats and gray tails (cinereicauda). They could represent subspecies of one species, the Variable Mountain-gem, as in Schuchman (1999 = HBW-5). Or the AOU could be right that they are two species, the Purple-throated Mountain-gem, Lampornis calolaemus, and the White-throated Mountain-gem, Lampornis castaneoventris (including cinereicauda). Alternatively, Stiles and Skutch (1989) and the IOC treat them as three allospecies, with Lampornis cinereicauda taking the name Gray-tailed Mountain-gem.
The DNA analysis revealed that these taxa are very closely related (García-Moreno et al., 2006), and is consistent with lumping all three forms into Variable Mountain-gem. However, although the ranges do meet, there is only limited evidence of hybridization, suggesting that they are best treated as three allospecies. That is the course followed here. In truth, there is insufficient data and a detailed study would be useful.
García-Moreno et al. (2006) found some evidence that White-bellied Mountain-gem, Lampornis hemileucus, does not belong with the other Lampornis but is closer to Panterpe. However, McGuire et al. (2009, 2014), which used a superset of the genes studied by García-Moreno et al., found the White-bellied Mountain-gem does in fact belong with other Lampornis.
Doubleday's Hummingbird, Chlorostilbon doubledayi, has been split from Broad-billed Hummingbird, Chlorostilbon latirostris. These two taxa have easily distinguished plumage, and in spite of the fact that their ranges abut one another, seem to be reciprocally monophyletic (García-Deras et al., 2008). In short, they appear to be distinct species, as treated by IOC, HBW-5 (Schuchmann, 1999), and Howell and Webb (1995), but not AOU.
Based on Feo et al. (2015) and the 56th AOU supplement, the Inagua Woodstar, Nesophlox lyrura, has been split from the Bahama Woodstar, Nesophlox evelynae.
Based on Gonzalez et al. (2011) and Navarro-Sigüenza, and Peterson (2004), the Wedge-tailed Sabrewing, Pampa (Campylopterus) curvipennis, has been split into Curve-winged Sabrewing, Pampa curvipennis (sister to Long-tailed Sabrewing, Pampa excellens) and Wedge-tailed Sabrewing, Pampa pampa, of the Yucatan.
The Dry Forest Sabrewing, Campylopterus calcirupicola, is recognized as a separate species related to the Gray-breasted Sabrewing, Campylopterus largipennis. See SACC #756 and Lopes et al. (2017). Lopes et al. and SACC #755 suggest there are additional species in this complex.
The Blossom-crown, Anthocephala floriceps, has been split into Santa Marta Blossomcrown, Anthocephala floriceps, and Tolima Blossomcrown, Anthocephala berlepschi. See Lozano-Jaramillo et al. (2014) and SACC proposal #654 (under consideration).
The Plovercrest, Stephanoxis lalandi, has been split into Purple-crowned Plovercrest, Stephanoxis loddigesii, and Green-crowned Plovercrest, Stephanoxis lalandi. See Cavarzere et al. (2014) and SACC proposal #664 (under consideration).
The Violet-crowned Woodnymph, Thalurania colombica, and Green-crowned Woodnymph, Thalurania fannyi, have been merged into Crowned Woodnymph, Thalurania colombica. See SACC proposal #558. Note that AOU's NACC has not acted on this yet.
The Streamertail, Trochilus polytmus, was split into Red-billed Streamertail, Trochilus polytmus, and Black-billed Streamertail, Trochilus scitulus. The AOU maintains these as one species, but Gill et al. (1973) provides evidence of a narrow hybrid zone. Accordingly, I've decided to follow the IOC and HBW-5 (Schuchmann, 1999) on this one.
Finally, the Blue-vented Hummingbird, Saucerottia hoffmanni, of Nicaragua and Costa Rica, has been split from the Steely-vented Hummingbird, Saucerottia saucerottei.
All this means there are seven non-AOU species in the list: Mexican Hermit, Gray-tailed Mountain-gem, Wedge-tailed Sabrewing, Doubleday's Hummingbird, Curve-winged Sabrewing, Black-billed Streamertail, and Blue-vented Hummingbird.