Thraupid Group
We've now reached the last set of families, the Mitrospingidae, Cardinalidae, and Thraupidae. It might make sense to to treat them all as subfamilies of an expanded Thraupidae. The genetic distance between them is small. Barker et al. (2013) estimate they have a common within the last 13 million years. In comparison, most families represent separate lineages stretching back 20 million years or more. Moreover, it's difficult to distinguish whether birds are cardinal-grosbeaks or tanagers (e.g., the Piranga tanagers, the saltators, the Paroaria cardinals, all of Mitrospingidae) suggests that it might be better to reduce both Mitrospingidae and Cardinalidae to subfamilies of Thraupidae.
Moreover, I am not the first to suggest downgrading them. Sibley and Monroe (1993) also noted the close relationship and went much further in combining families, treating the tanagers and cardinals as tribes within a broad subfamily Emberizinae which is equivalent to what is ranked here as the epifamily Icteroidae.
Mitrospingidae: Mitrospingus and allies Barker et al., 2013
3 genera, 4 species Not HBW Family
Although they are generally considered tanagers, there have been several papers indicating that Lamprospiza and Mitrospingus are not tanagers. Burns (1997) already found they lie outside the tanagers, and perhaps sister to Chlorospingus, now known to be a sparrow. Taxon sampling outside the tanagers was sparse, so Burns could not accurately place them. Yuri and Mindell (2002) provided additional evidence that they were not tanagers. The same was true of Burns et al. (2003). Klicka et al. (2007) had good sampling of all the relevant groups. They found Mitrospingus sister to the Emberizid group.
A very comprehensive study including them is Barker et al. (2013). Their results suggest that the two genera belong to the Thraupid group, and that Orthogonys belongs near them. Barker et al. established the family name Mitrospingidae for these three genera.
Mitrospingidae Species List
- Dusky-faced Tanager, Mitrospingus cassinii
- Olive-backed Tanager, Mitrospingus oleagineus
- Red-billed Pied Tanager, Lamprospiza melanoleuca
- Olive-green Tanager, Orthogonys chloricterus
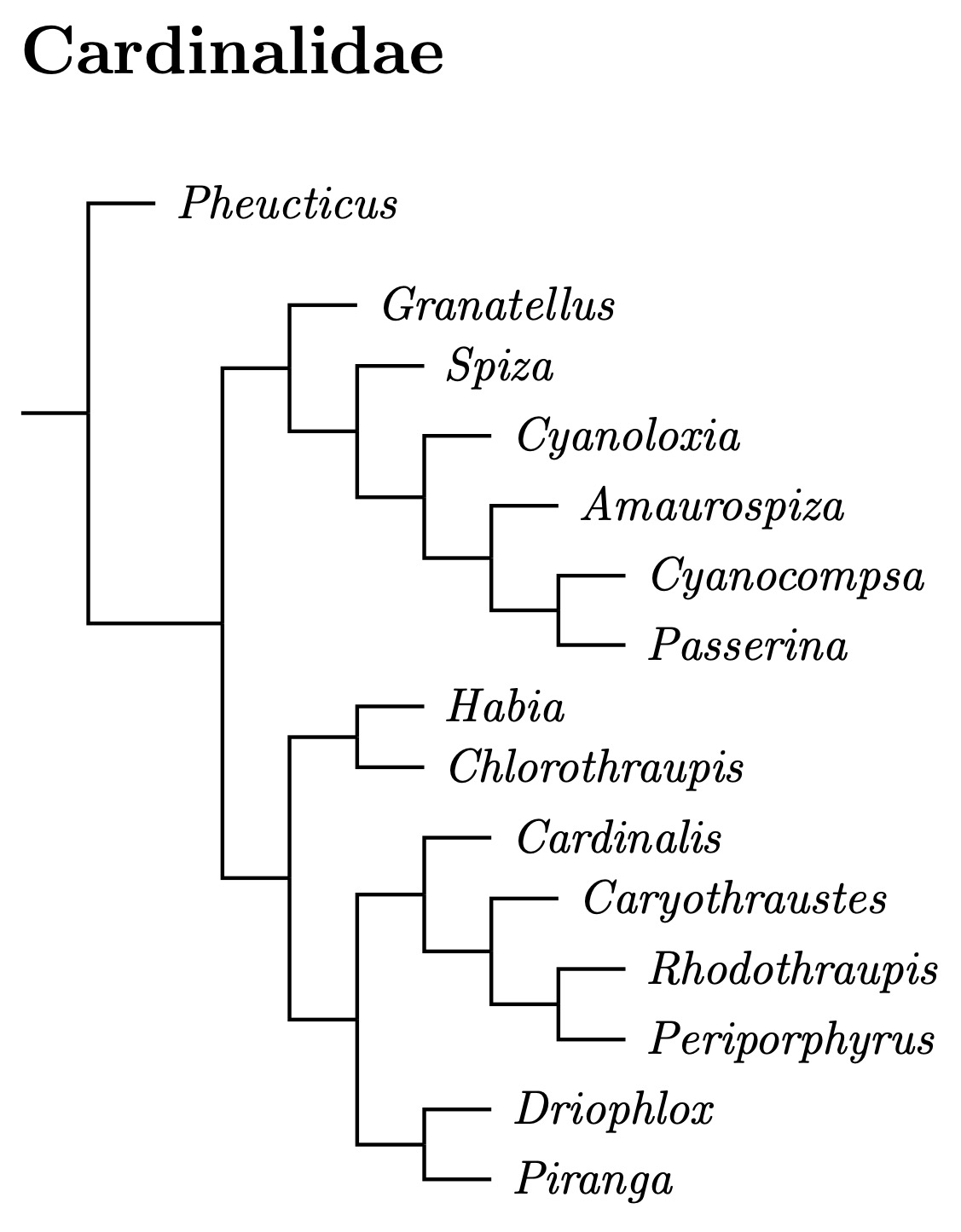
Cardinalidae: Cardinals, Grosbeaks Ridgway, 1901
15 genera, 56 species HBW-16
 |
| Click for Cardinalidae tree |
|---|
I had previously used Klicka et al. (2007) to reorganize the Cardinalidae. As a result, the saltators, Parkerthraustes, and Porphyrospiza joined the tanagers. In return, the cardinals got Piranga, Habia, and Chlorothraupis from the tanagers. They also gained the Granatellus chats from the warblers and the Amaurospiza blue seedeaters that are sometimes considered sparrows, and sometimes considered tanagers.
2024 Rearrangments
The major change is making the Pheucticus grosbeaks the basal group. The remaining Cardinalidae divide into two clades that, except for Pheucticus, are the same as before at the generic level. I flipped the order because the size of the Granatellus now contains fewer species.
Although the composition of the Cardinalidae remains the same, I've rearranged them some based on Scott's (2022) MS thesis, which uses ultra-conserved elements from DNA. I've rearranged the Cardinalidae, mostly using his Figure 1.1, most of which has 100% support.
The Habia Problem
I originally followed Klicka et al.'s suggestions of folding Chlorothraupis into Habia. However, the new phylogeny from Scott (2022) makes this untenable, as the expanded Habia consists of two separated parts. Fortunately, Scott, Chesser, Unitt, and Burns (2024) established the new genus name Driophlox, with type gutturalis (Sooty Ant-Tanager). This allows a more sensible division of these species.
With the new genus name in hand, we can solve the Habia problem by restricting Habia to its type, rubica (Red-crowned Ant-Tanager), restoring the former Chlorothraupis tanagers to Chlorothraupis, and putting the remaining four species in Driophlox. Then Driophlox is sister to Piranga and the Habia-Chlorothraupis group is sister to a clade consisting of Driophlox, Piranga, and 7 other species.
Following IOC and Ridgely (Ridgely and Greenfield, 2001; Ridgely and Tudor, 2009), the Olive Tanager, Chlorothraupis frenata, has been split from from Carmiol's Tanager, Chlorothraupis carmioli. This has also been followed by the SACC, in SACC proposal #950. However, instead of using Olive Tanager as the English name of Chlorothraupis frenata, they changed it to Yellow-lored Tanager at the same time. This avoids confusion with C. olivacea.
The genus Driophlox consists of:
- Red-throated Ant-Tanager, Driophlox fuscicauda
- Black-cheeked Ant-Tanager, Driophlox atrimaxillaris
- Sooty Ant-Tanager, Driophlox gutturalis
- Crested Ant-Tanager, Driophlox cristata
Moreover, Chlorothraupis is restored as:
- Ochre-breasted Tanager, Chlorothraupis stolzmanni
- Carmiol's Tanager, Chlorothraupis carmioli
- Lemon-spectacled Tanager, Chlorothraupis olivacea
- Yellow-lored Tanager, Chlorothraupis frenata
Older Changes with Updates
The order within Pheucticus was orginally based on Pulgarín-R et al. (2013). This was slightly modified using Scott (2022). Pulgarín-R et al. also found that two of the yellow grosbeaks were not monophyletic. The stronger case for a split is the Orange-colored Grosbeak, Pheucticus aurantiacus, which I have split from the (Mexican) Yellow Grosbeak, Pheucticus chrysopeplus. The Orange-colored Grosbeak is monotypic, distinctly orange, and can be found in Chiapis, Mexico and Guatemala.
There is a weaker case for splitting Pheucticus uropygialis from the Black-backed Grosbeak, Pheucticus aureoventris (monotypic). There's not an identification problem. The rump is variously yellow or mottled in the uropygialis group, black in aureoventris. However, the genetic distance is much smaller and there are questions about whether crissalis (part of the uropygialis group) interbreeds with chrysogaster in Ecuador. It could be that this complex is best treated as one species rather than three. For the present, I leave it at the conventional two.
Based on Bryson et al. (2014), I had previously moved most of Cyanocompsa into Cyanoloxia. However, I put the Blue Bunting in Passerina.
After considering Scott (2022) and the various species in Passerina, I've revived Cyanocompsa for the Blue Bunting, Cyanocompsa parellina, and the Turquoise-fronted Bunting, Cyanocompsa indigotica. They are sister to Passerina, but are distant enough genetically and different enough from Passerina to have their own genus. I had previously split the Turquoise-fronted Bunting, Cyanocompsa indigotica, of western Mexico from Blue Bunting, Cyanocompsa parellina, based on Bryson et al., (2014).
Within the other blue cardinalids, I've followed Bryson et al. (2014) by splitting Ecuadorian Seedeater, Amaurospiza aequatorialis, from Blue Seedeater, Amaurospiza concolor (which now takes the name Cabanis's Seedeater), and also splitting the Amazon Grosbeak, Cyanoloxia rothschildii, from Blue-black Grosbeak, Cyanoloxia cyanoides. The latter gives me an armchair tick as I have seen Amazon Grosbeak at Alta Floresta (Brazil) and Blue-black in the Canal Zone (Panama).
Klicka et al. (2001), Klicka et al. (2007), and Scott (2022) found that Lazuli Bunting, Passerina amoena, is sister to the Blue Grosbeak, P. caerulea, rather than to the Indigo Bunting, P. cyanea. This is apparently an artifact due to a combination of recent divergence and relatively large populations. In such cases, a longer time is necessary for all genes to fully reflect the species tree.
This has been studied in detail by Carling and Brumfield (2008), who found that the Lazuli and Indigo Buntings are sister species. As is well-known, they do hybridize in the contact zones of the western US. Carling and Brumfield (2009) and Carling et al. (2010) have also studied this phenomenon. Among other things, their further studies support the classification of Lazuli and Indigo Buntings as distinct and monotypic species. I still think this makes sense.
The arrangement of Piranga is based on Manthey et al. (2016). It agrees with the analysis of Scott (2022). The Hepatic Tanger has been split into four species: Northern Hepatic-Tanager / Hepatic Tanager, Piranga hepatica, Highland Hepatic-Tanager / Tooth-billed Tanager, Piranga lutea, Blood-red Tanager, Piranga haemalea, and Lowland Hepatic-Tanager / Red Tanager, Piranga flava, These have frequently been considered distinct species and some are so treated in many regional guidebooks. Burns (1998) found genetic differences between the three in the range typical of species, not subspecies. Manthey et al. (2016) clarified the position of Piranga haemalea.
Cardinalidae Species List
- Black-headed Grosbeak, Pheucticus melanocephalus
- Rose-breasted Grosbeak, Pheucticus ludovicianus
- Yellow Grosbeak / Mexican Yellow Grosbeak, Pheucticus chrysopeplus
- Orange-colored Grosbeak, Pheucticus aurantiacus
- Black-thighed Grosbeak, Pheucticus tibialis
- Golden Grosbeak / Southern Yellow Grosbeak, Pheucticus chrysogaster
- Black-backed Grosbeak, Pheucticus aureoventris
- Rose-breasted Chat, Granatellus pelzelni
- Red-breasted Chat, Granatellus venustus
- Gray-throated Chat, Granatellus sallaei
- Dickcissel, Spiza americana
- Blue-black Grosbeak, Cyanoloxia cyanoides
- Amazonian Grosbeak, Cyanoloxia rothschildii
- Ultramarine Grosbeak, Cyanoloxia brissonii
- Glaucous-blue Grosbeak, Cyanoloxia glaucocaerulea
- Cabanis's Seedeater, Amaurospiza concolor
- Ecuadorian Seedeater, Amaurospiza aequatorialis
- Carrizal Seedeater, Amaurospiza carrizalensis
- Blackish-blue Seedeater, Amaurospiza moesta
- Turquoise-fronted Bunting, Cyanocompsa indigotica
- Blue Bunting, Cyanocompsa parellina
- Blue Grosbeak, Passerina caerulea
- Lazuli Bunting, Passerina amoena
- Indigo Bunting, Passerina cyanea
- Rose-bellied Bunting, Passerina rositae
- Orange-breasted Bunting, Passerina leclancherii
- Varied Bunting, Passerina versicolor
- Painted Bunting, Passerina ciris
- Red-crowned Ant-Tanager, Habia rubica
- Ochre-breasted Tanager, Chlorothraupis stolzmanni
- Carmiol's Tanager, Chlorothraupis carmioli
- Lemon-spectacled Tanager, Chlorothraupis olivacea
- Yellow-lored Tanager, Chlorothraupis frenata
- Vermilion Cardinal, Cardinalis phoeniceus
- Northern Cardinal, Cardinalis cardinalis
- Pyrrhuloxia, Cardinalis sinuatus
- Black-faced Grosbeak, Caryothraustes poliogaster
- Yellow-green Grosbeak, Caryothraustes canadensis
- Crimson-collared Grosbeak, Rhodothraupis celaeno
- Red-and-black Grosbeak, Periporphyrus erythromelas
- Red-throated Ant-Tanager, Driophlox fuscicauda
- Black-cheeked Ant-Tanager, Driophlox atrimaxillaris
- Sooty Ant-Tanager, Driophlox gutturalis
- Crested Ant-Tanager, Driophlox cristata
- Red-headed Tanager, Piranga erythrocephala
- White-winged Tanager, Piranga leucoptera
- Red-hooded Tanager, Piranga rubriceps
- Rose-throated Tanager, Piranga roseogularis
- Scarlet Tanager, Piranga olivacea
- Western Tanager, Piranga ludoviciana
- Flame-colored Tanager, Piranga bidentata
- Summer Tanager, Piranga rubra
- Northern Hepatic-Tanager / Hepatic Tanager, Piranga hepatica
- Highland Hepatic-Tanager / Tooth-billed Tanager, Piranga lutea
- Blood-red Tanager, Piranga haemalea
- Lowland Hepatic-Tanager / Red Tanager, Piranga flava
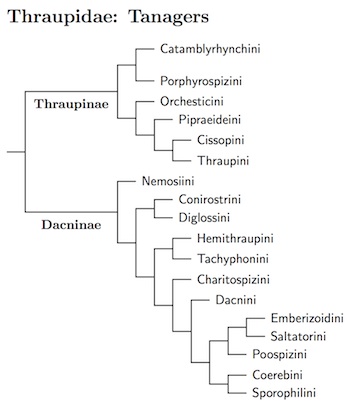
Thraupidae: Tanagers Cabanis, 1847
95 genera, 385 species HBW-16
 |
| Thraupidae family tree |
|---|
We finally reach the last family on the list! The tanagers have been the subject of an extensive ongoing revision. Kevin Burns and his co-workers have spent the last twenty years studying the tanagers. E.g., Burns (1997, 1998); Burns et al. (2002, 2003); Burns and Naoki (2004); Burns and Racicot (2009); Mauck and Burns (2009); Mallarinoa et al. (2012); Sedano and Burns (2010); Barker et al. (2013); Mason and Burns (2013); Ryan et al. (2013); Shultz and Burns (2013). This work has culminated in six-gene study Burns et al. (2014), which is the primary source for the TiF phylogeny of the tanagers.
In recent years, the tanagers lost the euphonias and chlorophonias to the finches, Habia and Piranga (including all of the North American tanagers) to the cardinals, the Mitrospingidae, and the Phaenicophilidae. However, the tanagers have also gained many species. The tanager-finches were historically often considered sparrows mostly end up in the tanagers. The tanagers also gained the saltators, some other cardinals, and the bananaquit.
The result of this shifting genera is a very large, very heterogeneous tanager family. This is especially remarkable as the tanagers are a relatively recent radiation. There is a basal division into two parts, Thraupinae and Dacninae. I have found it useful to further divide these into 18 tribes (15 of these are the “subfamilies” of Burns et al., 2014; 3 are further subdivisions of these).
Thraupidae Subfamilies
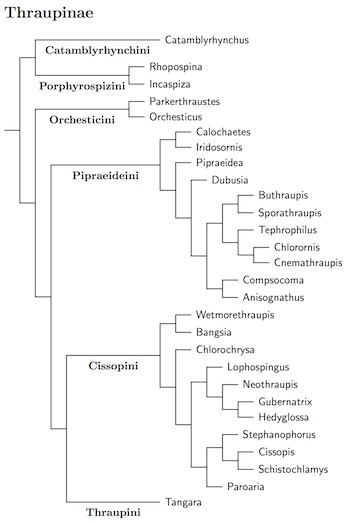
Thraupinae Cabanis, 1847
 |
| Thraupinae subfamily tree Click for Thraupinae species tree |
|---|
The Thraupinae are shown in more detail at the right. There are six groups: Catamblyrhynchini and Porphyrospizini (formerly Incaspizini), and the Orchesticini, Thaupini, Cissopini, Pipraeideini clade.
Catamblyrhynchini: Plushcap
This contains a single species, Plushcap. It's closest relatives appear to be the Porphyrospizini, but the relationship is rather distant.
Porphyrospizini: Inca-Finches
I had previously tentatively placed Incaspiza and its allies basally in Poospizini. However, Barker et al. (2013) was the first to sequence any of the Incaspiza, finding that the Great Inca-Finch grouped with the Blue Finch and Mourning Sierra Finch.
The Rhopospina are rather distant from the other sierra finches (Campanga et al., 2011, Klicka et al., 2007). Ridgely and Tudor (1989) long ago grouped the Band-tailed, Carbonated and Mourning Sierra Finches based on plumage. Burns et al. (2014) found that the Blue Finch (Porphyrospiza) also belongs in this clade. The display flights may also indicate a connection. Campagna et al. (2011) found that they are each other's closest relatives, and quite distant from any of the other sierra finches. This small clade takes the name Rhopospina (Cabanis 1851, type fruticeti) which has priority over Corydospiza (Sundevall 1872, type alaudina) and Porphyrospiza (PL Sclater and Salvin 1873, type caerulescens).
The rest of Thraupinae is in the main clade. This includes four successive branches: Orchesticini, Pipraeideini, Cissopini, and Thraupini.
Orchesticini: Grosbeak-Tanagers
This is a small group of two species, Parkerthraustes and Orchesticus that Burns et al. (2014) found to be basal to the main clade, and not particularly closely related to them. The relationships of these two species have been murky. Weir et al. (2009), using only cytochrome-b, found Parkerthraustes basal in Saltatorinae. Klicka et al. (2007), using more genes, found it sister to Chlorochrysa, but with weak support. Finally, the more comprehensive analysis of Barker et al. (2013) put it sister to Orchesticus with very strong support, and the two of them were weakly attached to the Hemithraupini clade.
Pipraeideini: Mountain-Tanagers
That brings us to Pipraeideini. These are almost all mountain-tanagers, birds of the Andes. Here again I use an informal name for the tribe as none have been established in the literature.
Based on the topology of Sedano and Burns (2010) and SACC decision #569 several of the genus boundaries have been changed. The Blue-and-yellow Tanager, formerly Thraupis bonariensis, is now in Pipraeidea. The genus Delothraupis (Chestnut-bellied Mountain-Tanager) has been merged into Dubusia. It is sister to the former Rufous-bellied Saltator, Saltator rufiventris, which is now called Rufous-bellied Mountain-Tanager, Pseudosaltator rufiventris due to SACC Proposal #722.
The genus Buthraupis has been dismembered, leaving only the Hooded Mountain-Tanager, Buthraupis montana. The Grass-green Tanager is now in the monotypic Chlorornis (Reichenbach, 1850), and the Masked Mountain-Tanager is in the monotypic Tephrophilus (Moore 1934), while the Black-chested and Golden-backed Mountain-Tanagers have been put in Cnemathraupis (Penard, 1919, type eximia). The Blue-capped Tanager, formerly Thraupis cyanocephalus, is now in the monotypic Sporathraupis (Bonaparte, 1850). The Blue-winged Mountain-Tanager, Anisognathus somptuosus, and Black-chinned Mountain-Tanager, Anisognathus notabilis, have been placed in genus Compsocoma (Cabanis, 1853, type somptuosa victorini).
Cissopini: Cardinal-Tanagers
Both Weir et al. (2009) and Sedano and Burns (2010) put Chlorochrysa in Cissopini, and we follow that here. Note that Klicka et al. (2007) found a somewhat different arrangement, with Chlorochrysa sister to Parkerthraustes, and the pair basal to several tanager tribes.
Klicka et al. (2014) found that the Yellow-green Bush-tanager, formerly Chlorospingus flavovirens, was a tanager, and not part of the sparrow genus Chlorospingus. However, they did not include enough tanagers in their analysis to determine its affinities. This has now been remedied by Avendaño et al. (2016), who found it is sister to the Blue-and-gold Tanager, Bangsia arcaei. As a result, the Yellow-green Bush-tanager is now Bangsia flavovirens.
The Paroria and Gubernatrix cardinals (often considered sparrows) are in Cissopini with several other tanagers and finches. The only DNA information available concerning Gubernatrix is from Barker et al. (2013). The species limits of Paroaria are a bit non-standard. Based on Dávalos and Porzecanski (2009), I've separated Masked Cardinal, Paroaria nigrogenis, from Red-capped Cardinal, P. gularis, and moved the subspecies cervicalis of eastern Boliva and NW Mato Grosso into Yellow-billed Cardinal, P. capitata. (An alternative would be to lump capitata with gularis as Red-capped Cardinal, as suggested many years ago by Hellmayr (1938)). Thus Yellow-billed Cardinal includes the subspecies cervicalis, capitata, and fuscipes. Based on Lopes and Gonzaga (2013), I have also split the Crimson-fronted Cardinal into the black-throated Xingu Cardinal, Paroria xinguensis and crimson-throated Araguaia Cardinal, Paroria baeri.
Thraupini: Tangara Tanagers
In the current phylogeny, Thraupini is the biggest group in Thraupinae. It consists solely of the genus Tangara. That means it includes some of the most attractive birds on the planet. You may be wondering why it's called Thraupini when the only genus present is Tangara. Most of the genus Thraupis, including the terminologically important type species (formerly Thraupis ornata) have been subsumed in Tangara as a result of Sedano and Burns (2010). Although Tangara has priority (by seniority) over Thraupis at the genus-level, an official ruling means that Thraupis has priority at the family (and tribal) level. Thus the tribe containing Tangara ornata is known as the Thraupini.
If you example the species-level tree, you'll see that Tangara contains two clades, which could legitimately be called Tangara and Thraupis. I've chosen to retain these as subgenera, but hope that the AOU will do the sensible thing and use these these as genus names. Arguably I should go ahead and use them here, but at present I prefer that one of the AOU committees take the lead.
A close examination of the species-level Thraupini tree also reveals the numbers 1-13 labelling most of the clades in Tangara. These indicate the numbered clades identified by Isler and Isler (1987) using traditional taxonomic methods. Except for clades 3 and 9, they match up precisely with the genetic data. The clade labelled “Th” consists of the Tangara species formerly placed in Thraupis.
The Black-headed Tanager had to take an alternate name, Tangara argentea, because the Azure-shouldered Tanager, formerly Thraupis cyanoptera, has first claim on Tangara cyanoptera.
Thraupinae Species List
Catamblyrhynchini: Plushcap Ridgway, 2014
- Plushcap, Catamblyrhynchus diadema
Porphyrospizini: Inca Finches and allies Burns et al., 2014
- Mourning Sierra Finch, Rhopospina fruticeti
- Blue Finch, Rhopospina caerulescens
- Band-tailed Sierra Finch, Rhopospina alaudina
- Carbonated Sierra Finch, Rhopospina carbonaria
- Great Inca-Finch, Incaspiza pulchra
- Rufous-backed Inca-Finch, Incaspiza personata
- Gray-winged Inca-Finch, Incaspiza ortizi
- Buff-bridled Inca-Finch, Incaspiza laeta
- Little Inca-Finch, Incaspiza watkinsi
Orchesticini: Grosbeak-Tanagers Burns et al., 2014
- Yellow-shouldered Grosbeak, Parkerthraustes humeralis
- Brown Tanager, Orchesticus abeillei
Pipraeideini: Mountain-Tanagers Informal
- Vermilion Tanager, Calochaetes coccineus
- Purplish-mantled Tanager, Iridosornis porphyrocephalus
- Yellow-throated Tanager, Iridosornis analis
- Golden-collared Tanager, Iridosornis jelskii
- Golden-crowned Tanager, Iridosornis rufivertex
- Yellow-scarfed Tanager, Iridosornis reinhardti
- Fawn-breasted Tanager, Pipraeidea melanonota
- Blue-and-yellow Tanager, Pipraeidea bonariensis
- Rufous-bellied Mountain-Tanager, Pseudosaltator rufiventris
- Buff-breasted Mountain-Tanager, Dubusia taeniata
- Chestnut-bellied Mountain-Tanager, Dubusia castaneoventris
- Hooded Mountain-Tanager, Buthraupis montana
- Blue-capped Tanager, Sporathraupis cyanocephala
- Masked Mountain-Tanager, Tephrophilus wetmorei
- Grass-green Tanager, Chlorornis riefferii
- Black-chested Mountain-Tanager, Cnemathraupis eximia
- Golden-backed Mountain-Tanager, Cnemathraupis aureodorsalis
- Blue-winged Mountain-Tanager, Compsocoma somptuosa
- Black-chinned Mountain-Tanager, Compsocoma notabilis
- Black-cheeked Mountain-Tanager / Santa Marta Mountain-Tanager, Anisognathus melanogenys
- Lacrimose Mountain-Tanager, Anisognathus lacrymosus
- Scarlet-bellied Mountain-Tanager, Anisognathus igniventris
Cissopini: Cardinal-Tanagers Sundevall, 1872
- Orange-throated Tanager, Wetmorethraupis sterrhopteron
- Blue-and-gold Tanager, Bangsia arcaei
- Yellow-green Bush-tanager, Bangsia flavovirens
- Black-and-gold Tanager, Bangsia melanochlamys
- Golden-chested Tanager, Bangsia rothschildi
- Moss-backed Tanager, Bangsia edwardsi
- Gold-ringed Tanager, Bangsia aureocincta
- Glistening-green Tanager, Chlorochrysa phoenicotis
- Multicolored Tanager, Chlorochrysa nitidissima
- Orange-eared Tanager, Chlorochrysa calliparaea
- Black-crested Finch, Lophospingus pusillus
- Gray-crested Finch, Lophospingus griseocristatus
- White-banded Tanager / Shrike-like Tanager, Neothraupis fasciata
- Yellow Cardinal, Gubernatrix cristata
- Common Diuca-Finch, Hedyglossa diuca
- Diademed Tanager, Stephanophorus diadematus
- Magpie Tanager, Cissopis leverianus
- Black-faced Tanager, Schistochlamys melanopis
- Cinnamon Tanager, Schistochlamys ruficapillus
- Red-crested Cardinal, Paroaria coronata
- Red-cowled Cardinal, Paroaria dominicana
- Masked Cardinal, Paroaria nigrogenis
- Red-capped Cardinal, Paroaria gularis
- Yellow-billed Cardinal, Paroaria capitata
- Xingu Cardinal, Paroaria xinguensis
- Araguaia Cardinal, Paroaria baeri
Thraupini: Tangara Tanagers Cabanis, 1847
- Blue-and-black Tanager, Tangara vassorii
- Beryl-spangled Tanager, Tangara nigroviridis
- Green-naped Tanager, Tangara fucosa
- Spangle-cheeked Tanager, Tangara dowii
- Blue-browed Tanager, Tangara cyanotis
- Metallic-green Tanager, Tangara labradorides
- Rufous-cheeked Tanager, Tangara rufigenis
- Rufous-winged Tanager, Tangara lavinia
- Bay-headed Tanager, Tangara gyrola
- Golden-eared Tanager, Tangara chrysotis
- Saffron-crowned Tanager, Tangara xanthocephala
- Flame-faced Tanager, Tangara parzudakii
- Green-and-gold Tanager, Tangara schrankii
- Blue-whiskered Tanager, Tangara johannae
- Golden Tanager, Tangara arthus
- Emerald Tanager, Tangara florida
- Silver-throated Tanager, Tangara icterocephala
- Plain-colored Tanager, Tangara inornata
- Turquoise Tanager, Tangara mexicana
- Paradise Tanager, Tangara chilensis
- Opal-rumped Tanager, Tangara velia
- Opal-crowned Tanager, Tangara callophrys
- Green-headed Tanager, Tangara seledon
- Seven-colored Tanager, Tangara fastuosa
- Red-necked Tanager, Tangara cyanocephala
- Gilt-edged Tanager, Tangara cyanoventris
- Brassy-breasted Tanager, Tangara desmaresti
- Dotted Tanager, Tangara varia
- Rufous-throated Tanager, Tangara rufigula
- Spotted Tanager, Tangara punctata
- Speckled Tanager, Tangara guttata
- Yellow-bellied Tanager, Tangara xanthogastra
- Golden-naped Tanager, Tangara ruficervix
- Blue-gray Tanager, Tangara episcopus
- Sayaca Tanager, Tangara sayaca
- Glaucous Tanager, Tangara glaucocolpa
- Azure-shouldered Tanager, Tangara cyanoptera
- Yellow-winged Tanager, Tangara abbas
- Palm Tanager, Tangara palmarum
- Golden-chevroned Tanager, Tangara ornata
- Gray-and-gold Tanager, Tangara palmeri
- Azure-rumped Tanager / Cabanis's Tanager, Tangara cabanisi
- Black-headed Tanager, Tangara argentea
- Silvery Tanager / Silver-backed Tanager, Tangara viridicollis
- Sira Tanager, Tangara phillipsi
- Black-capped Tanager, Tangara heinei
- Green-throated Tanager / Straw-backed Tanager, Tangara argyrofenges
- Golden-hooded Tanager, Tangara larvata
- Masked Tanager, Tangara nigrocincta
- Blue-necked Tanager, Tangara cyanicollis
- Black-backed Tanager, Tangara peruviana
- Chestnut-backed Tanager, Tangara preciosa
- Green-capped Tanager, Tangara meyerdeschauenseei
- Scrub Tanager, Tangara vitriolina
- Burnished-buff Tanager, Tangara cayana
- Lesser Antillean Tanager, Tangara cucullata
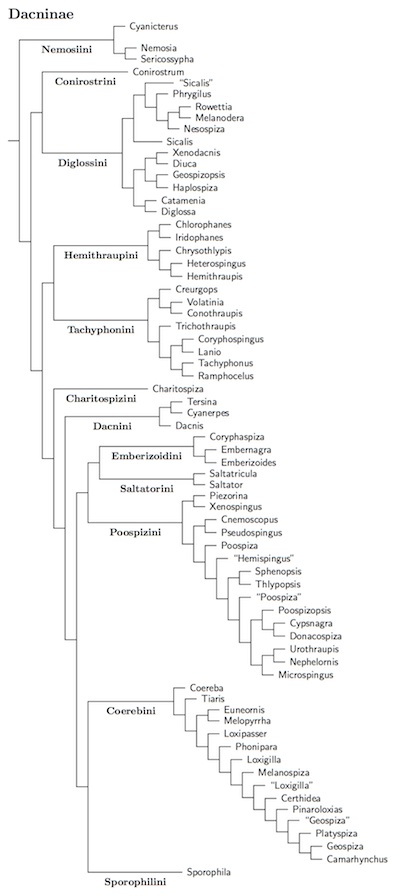
Dacninae Sundevall, 1836
 |
| Dacninae subfamily tree |
|---|
The rest of the Neotropical finches are part of Dacninae (a few remain outside the tanagers). This part of the tanagers is full of seedeaters, seedfinches, grass-finches, warbling finches, sierra finches, pileated finches, yellow finches, etc, etc, but not the brushfinches. It also contains the flower-piercers. The Dacninae does contain birds other than Neotropical finches. It includes the conebills, honeycreepers, and even some tanagers such as the striking Silver-beaked Tanager.
As with the Thraupinae, the arrangement here is based on Burns et al. (2014).
Nemosiini: Flocking-dwelling Tanagers
Barker et al. (2013) found that the previously untested Cyanicterus and Compsothraupis tanagers belong in this small clade which has 100% bootstrap support. There had been question about whether the similarity between Compsothraupis (Scarlet-throated Tanager) and Sericossypha (White-capped Tanager) was convergence or inheritance. Storer argued that it was convergence, a view endorsed by Ridgely and Tudor (1989). The jury is now in. Barker et al. found they are sister species and so I have merged Compsothraupis (Richmond 1915) into Sericossypha (Lesson 1844).
Conirostrini: Conebills
The conebills are in their own clade, Conirostrini. There is general consensus that the Conirostrini are sister to Diglossini. They could be treated as a single tribe under the name Diglossini. Since the conebills are distinctive and the division is relatively deep (Weir et al., 2009), I leave them separate.
The Giant Conebill is usually placed in the genus Oremanes as Oremanes fraseri (Sclater 1860). However, it turns out to be nested in Conirostrum, and so has been changed. This creates a bit of a nomenclatural complication as fraseri is preoccupied by Conirostrum cinereum fraseri (Sclater 1859). The next oldest available name for the Giant Conebill appears to be binghami (Chapman 1919).
Diglossini: Highland Tanagers
That brings us to the last group, the Diglossini finches. The arrangement here draws heavily on Campagna et al. (2011) and Mauck and Burns (2009). Barker et al. (2013) allowed me compeletely resolve the tree in a reasonable way.
Diglossini contains two clades. The first consists of yellow finches. Ryan et al. (2013) have eliminated the guesswork here. They found that Rowettia and Melanodera were each other's closest relatives, with Nesospiza sister to the pair. The whole clade is sister to the true Phrygilus sierra finches. Although this was the most likely arrangement, there was some support for Melanodura and Nesospingus as sisters, with Rowettia sister to the pair. Either way, the Atlantic islands of Tristan de Cuha and Gough have twice been colonized by South American finches.
The arrangement within Sicalis is based on Ryan et al. (2013), who included all but one species in their analysis. Their analysis also raised doubts about whether the Stripe-tailed Yellow-Finch belongs in Sicalis. In fact, most genes placed it outside Sicalis, but there was no agreement on whether it is closer to the true Phrygilus, basal to both, or even closer to the Melanodera/Rowettia/Nesospiza clade. For now, it is placed in the monotypic Pseudochloris (Sharpe 1888).
The Monte Yellow-Finch, Sicalis mendozae, has been split from Greenish Yellow-Finch, Sicalis olivascens. See Areta et al. (2012) and the discussion of SACC proposal 539.
The other clade includes the flowerpiercers, Tit-like Dacnis, and various gray finches. It's a bit unclear exactly where Catemenia fits. I'm following Barker et al. (2013) who place it sister to Diglossa, although Campagna et al. (2011) give a slightly different result (both are strongly supported). The two Idiopsar sierra finches have also been removed from Phrygilus. It might be better to put them in their own genus, sister to Idiopsar, but no genus name is available for them. The Geospizopsis sierra finches were formerly part of Phrygilus. Campagna et al. found them close to Haplospiza, and presumably Acanthidops. Note that all of the flower-piercers are in Diglossa, including those formerly placed in Diglossopis (see Mauck and Burns, 2009).
There may be a case for merging Acanthidops into Haplospiza. Weir et al. (2009) found that Acanthidops was more closely related to H. unicolor than to H. rustica. This is based on a single gene (cytochrome-b), so it is probably a bit premature to make the change.
Hemithraupini: Yellow-and-black Tanagers
All the members of this colorful group have been sequenced (see Barker et al., 2013; Burns et al., 2014; Weir et al. 2009). The Scarlet-and-white Tanager is sometimes put in a separate genus, Erythrothlypis. Weir et al. found it sister to the Black-and-yellow Tanager, so I leave them both in Chrysothlypis.
Tachyphonini: Ornamented Tanagers
Besides the complete analysis of Burns et al. (2014), other relevant papers include Burns and Racicot (2009), sorted out a big chunk of the Tachyphonini (which could equally be called Ramphocelini as both were established by Bonaparte in the same publication). Barker et al. (2013) found strong support for this clade, with mediocre support for including Volatinia, Creurgops, and Conothraupis as basal members. Note however that the two Creurgops show up next to the Poospzini in Weir et al. (2009), and in one of the two analyses in Burns et al. (2003).
The genus Tachyphonus was found to be paraphyletic, with part more closely related to Lanio, and part more closely related to Ramphocelus. The monotypic genus Eucometis has been merged into Trichothraupis, where it joins a couple of ex-Tachyphonus. Another monotypic genus, Rhodospingus, has joined Lanio, as has part of Tachyphonus.
Burns and Racicot did not consider whether or not the Lemon-rumped Tanager (R. flammigerus icteronotus) and Flame-rumped Tanager (R. f. flammigerus) should be split. They did not have samples from flammigerus, but they did find substantial genetic diversity within icteronotus. There is a hybrid zone in Colombia, which has been examined by Morales-Rozo et al. (2014). I don't see the case for a split.
Charitospizini: Coal-crested Finch
This tribe contains a single species, Coal-crested Finch. Barker et al. (2013) found that the Coal-crested Finch, Charitospiza eucosma did not belong with any of the other tribes. They put it in the middle of the Dacninae, but the node connecting it had quite low support (about 15% bootstrap). Burns et al. (2014) placed it on a separate branch, basal to a clade containing Dacini and the rest of the Dacinae.
Dacnini: Blue Tanagers
Dacinini includes the dacnises, the Cyanerpes honeycreepers, and the Swallow Tanager. The order within Dacnis is based on Burns et al. (2014).
Based on IOC and Ridgely and Greenfield (2001), Yellow-tufted Dacnis, Dacnis egregia (inc. aequatorialis) has been split from Black-faced Dacnis, Dacnis lineata.
Emberizoidini: Grassland Tanagers
This clade includes the grass and pampa finches. Embernagra and Emberizoides have grouped together in several studies. Barker et al. (2013) found that Coryphaspiza belongs with them to form a strongly supported clade. They also found some support for grouping them with the saltators, which is also supported by Burns et al. (2014).
Saltatorini: Saltators
Saltatricula is not particularly close to the Saltator saltators. The Black-throated Saltator has been moved to Saltatricula. The Rufous-bellied Saltator, “Saltator” rufiventris, is not really a saltator, and so has been moved to Dubusia in the Pipraeideini, as shown on the Thraupinae diagram. Finally, Pitylus has been merged into Saltator. Chaves et al. (2013) was the first analysis to include all of the saltator species. The order of the saltators reflects the results in Burns et al. (2014).
The Grayish Saltator has been split into three species based on Chaves et al. (2013). The three species are Middle American Saltator, Saltator grandis (all Middle American races); Plumbeous Saltator, Saltator plumbeus (presumed to include plumbeus and brewsteri); and Grayish Saltator, Saltator coerulescens (all other races). There may be more than three species here. In particular, plumage suggests that Brewster's may need to be separated from Plumbeous, but there is currently no genetic data.
Poospizini: Warbler-Tanagers
Poospizini is a big Neotropical finch group. The name Nephelornithini is equally applicable.
Until recently, the phylogentic picture here is clouded by the fact that neither Hemispingus nor Poospiza is monophyletic (see García-Moreno et al., 2001; Lougheed et al., 2000). Further, part of Phrygilus seems to be here too (see Klicka et al., 2007). The situation has improved with the appearance Shultz and Burns (2013), which has sorted out the various Hemispingus and Poospiza species. The result is that both Poospiza and Hemispingus are dismembered, with Poospiza spread across four different genera and Hemispingus across six. The generic name Hemispingus may disapper as a result.
The Gray-hooded Bush-Tanager (Cnemoscopus) is sister to a pair of hemispinguses, the Black-headed and Drab Hemispinguses. These are moved to the genus Pseudospingus (Berlepsch and Stolzmann 1896, type xanthophthalmus).
Although early indications were that the Cochabamba and Tucuman Mountain-Finches were separated from the other Poospiza, the more complete taxon sampling in Shultz and Burns (2013) shows that they are actually embedded in the main body of Poospiza. This means that the genus Compsospiza (Berlepsch 1893, type garleppi) must again be submerged in Poospiza (Cabanis 1847, type nigrorufa). Shultz and Burns suggest that their genetic results would support splitting Black-and-chestnut Warbling-Finch, P. whitii, from Black-and-rufous Warbling-Finch, Poospiza nigrorufa. The additional evidence from Jordan et al. (2017) is sufficient to justify the split. See also SACC Proposal 753.
The true Poospiza group is sister to the remaining Poospizini.
Based on García-Moreno et al. (2001, 2003), White-browed Hemispingus, Kleinothraupis auricularis, is split from Black-capped Hemispingus, Kleinothraupis atropileus. Although García-Moreno et al. were able to identify the main clades in the former Hemispingus, they weren't able to discern their relationships. They had trouble with the Gray-capped Hemispingus, Kleinothraupis reyi, which is the basal member of this Kleinothraupis group. The name Kleinothraupis was created by Burns et al. (2016) as there was not an available name for the group.
The Piura Hemispingus, Sphenopsis piurae, including macrophrys, is split from Black-eared Hemispingus, Sphenopsis melanotis. These two and frontalis form a small clade for which the name Sphenopsis (PL Sclater 1862, type frontalis) is available. Some authorities split Western Hemispingus (S. ochraceus) from Black-eared. What litte genetic data is available does not support this (García-Moreno et al., 2001).
Sphenopsis is sister to Thlypopsis, which has absorbed Pyrrhocoma and the Superciliaried Hemispingus. This creates a couple of nomenclatural problems which cannot be avoided. The first is that two of these species have the specific epithet ruficeps, the Chestnut-headed and Rust-and-yellow Tanagers. The Rust-and-yellow has priority (d'Orbigny and Lafresnay, 1837) over the Chestnut-headed (Strickland, 1844). However, there seem to be no junior names for the Chestnut-headed. Burns et al. (2016) have now created the name Thlypopsis pyrrhocoma for it. The other problem is that this group now contains the type species of three genera (Hemispingus, Pyrrhocoma, and Thlypopsis). All were named by Cabanis in the same publication in 1851, and none have priority over the others. A first reviser action is needed to resolve this. As most of these species have been in Thlypopsis, I will use that until the nomenclatural issue is resolved.
Another stray Poospiza is in a clade by itself which was named Castanozoster by Burns et al. (2016). Thus I refer to the Bay-chested Warbling-Finch as Castanozoster thoracicus. Two more Poospiza take the name Poospizopsis (Berlepsch 1893, type caesar). They are sister to Cypsnagra and Donacospiza. These three genera together are sister to Urothraupis, Nephelornis, and a collection of more Poospiza and a Hemispingus. The name Microspingus (Taczanowski 1874, type trifasciatus) is available for this group, which is at the end of the Poospizini.
Beldsoe (1988) showed that Urothraupis belongs with the tanagers, but gives little guidance as to where in the tanagers. It has sometimes been considered close to the Chlorospingus bush tanagers, but that is incorrect since they are not tanagers!
Coeribini: Dome-nesting Tanagers
The Coerebini are not the old honeycreeper family, but are a clade of mainly island species including West Indian “quits” and bullfinches, and Darwin's Finches. The term Tholospiza (dome finch) was introduced by Burns et al. (2002) to avoid confusion with the old honeycreeper family (Coeribidae), but Coeribini has priority.
I've changed several of the generic boundaries in Coeribini to reflect the genetic tree found by Burns et al. (2014). The Barker et al. (2013) version is similar. The Puerto Rican and Greater Antillean Bullfinches move from Loxigilla to join the Cuban Bullfinch in Melopyrrha. Only the Yellow-faced Grassquit remains in Tiaris. The rest of Tiaris moves to Loxigilla, as does the St. Lucia Black Finch (formerly the only member of Melanospiza). The Dull-colored and Sooty Grassquits are in a clade of their own, named Asemospiza by Burns et al. (2016).
The Grand Cayman Bullfinch, Melopyrrha taylori, has been split from the Cuban Bullfinch, Melopyrrha nigra (see Garrido et al., 2014). They differ in plumage, morphology, size, and song.
Darwin's finches are quite closely related and introgression makes it hard to discern the actual relationships between them. Previous versions wereh based on Petren et al. (2005), Tonnis et al. (2005), and the discussion in Grant and Grant (2008). After considering Farrington et al. (2014) and Lamichhaney et al. (2015), I have rearranged Darwin's Finches. I also merged Camarhynchus into Geospiza, and split some taxa.
- Sharp-beaked Ground-Finch, Geospiza difficilis has been split into
- Sharpe's Ground-Finch, Geospiza difficilis (including debilirostris),
- Vampire Finch, Geospiza septentrionalis, and
- Sharp-beaked Ground-Finch, Geospiza acutirostris
- Large Cactus-Finch, Geospiza conirostris, has been split into
- Large-billed Cactus-Finch, Geospiza conirostris (possibily including darwini), and
- Genovesa Cactus-Finch, Geospiza propinqua
This leads to a net gain of 3 species. However, there is a case to be made for lumping conirostris into magnirostris, propinqua into scandens, and acutirostris into fortis. If the distribution table below is correct, each pair is allopatric, and the divergence times fairly short. The three pairs occupy the last 6 rows of the table. The full tree is shown in the species list.
| Distribution of Darwin's Finches in the Galápagos Islands | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Sey | Bal | Isa | Fer | Stg | Rab | Pnz | SCrz | Dap | SFe | SCrst | Esp | Flo | Gen | Mar | Pnt | Dar | Wol |
| C. fusca | B | B | B | E | B | B | B | B | B | |||||||||
| C. olivacea | B | B | B | B | B | B | ||||||||||||
| Pl. crassirostris | B | B | B | (E) | E | B | (E) | B | B | B | B | |||||||
| G. difficilis | (E) | B | B | E | (E) | E | B | |||||||||||
| G. psittacula | B | B | B | (E) | E | B | B | B | B | B | ||||||||
| G. pauper | B | |||||||||||||||||
| G. parvula | B | B | B | (E) | B | B | B | B | B | |||||||||
| G. heliobates | B | E | ||||||||||||||||
| G. pallida | B | (B) | B | B | B | B | ||||||||||||
| G. septentrionalis | B | B | ||||||||||||||||
| G. fuliginosa | B | B | B | B | B | B | B | B | B | B | B | B | B | B | B | |||
| G. fortis | B | B | B | B | B | B | B | B | B | B | B | B | B | B | ||||
| G. acutirostris | B | |||||||||||||||||
| G. magnirostris | B | B | B | B | B | B | B | B | E | E | B | B | B | B | B | |||
| G. conirostris | B | |||||||||||||||||
| G. scandens | B | B | B | B | B | (B) | B | B | B | B | B | B | B | |||||
| G. propinqua | B | |||||||||||||||||
|
In the table, B means breeding and E means previously breeding, but now
locally extinct, while parenthese indicate uncertainty. Location information
is primarily from Grant and Grant (2008). They appear to consider
darwini, which is on Darwin and maybe Wolf, a subspecies of
magnirostris rather than conirostris. Eighteen of the Galápagos Islands are listed. They are: Seymour, Baltra (South Seymour), Isabela (Albemarle), Fernandina (Narborough), Santiago (James), Rábida (Jervis), Pinzón (Duncan), Santa Cruz (Indefatigable), Daphne, Santa Fe (Barrington) (Barrington), San Cristóbal (Chatham), Espanñola (Hood), Floreana (Charles), Genovesa (Tower), Marchena (Bindloe), Pinta, Darwin (Culpepper), and Wolf (Wenham). | ||||||||||||||||||
Sporophilini: Seedeaters
Mason and Burns (2013) showed that both Dolospingus and Oryzoborus are embedded in Sporophila, as expected. As a result, both Dolospingus and Oryzoborus have been merged into Sporophila. The arrangement of the Sporophilini now follows Mason et al. (2016).
Following an SACC decision, Capped Seedeater, Sporophila bouvreuil, has been split into Pearly-bellied Seedeaster, Sporophila pileata, and Copper Seedeater, Sporophila bouvreuil. See Machado and Silveira (2010; 2011). Further, the yellow-billed Tropeiro Seedeater, Sporophila beltoni, has been split from the dark-billed Plumbeous Seedeater, Sporophila plumbea (Repenning and Fontana, 2013).
Based on Mason et al. (2016), the White-collared Seedeater has been split into Cinnamon-rumped Seedeater, Sporophila torqueola (Western and Central Mexico) and Morellet's Seedeater / White-collared Seedeater, Sporophila morelleti (the rest of the range). These are not even sister taxa.
Dacninae Species List
Nemosiini: Flock-dwelling Tanagers Bonaparte, 1854
- Hooded Tanager, Nemosia pileata
Click for Dacninae tree
part I: Nemosiini —
Tachyphonini - Cherry-throated Tanager, Nemosia rourei
- Blue-backed Tanager, Cyanicterus cyanicterus
- White-capped Tanager, Sericossypha albocristata
- Scarlet-throated Tanager, Sericossypha loricata
Conirostrini: Conebills Edwards, 1986
- Bicolored Conebill, Conirostrum bicolor
- Pearly-breasted Conebill, Conirostrum margaritae
- Chestnut-vented Conebill, Conirostrum speciosum
- White-eared Conebill, Conirostrum leucogenys
- Capped Conebill, Conirostrum albifrons
- Giant Conebill, Conirostrum binghami
- Blue-backed Conebill, Conirostrum sitticolor
- White-browed Conebill, Conirostrum ferrugineiventre
- Tamarugo Conebill, Conirostrum tamarugense
- Rufous-browed Conebill, Conirostrum rufum
- Cinereous Conebill, Conirostrum cinereum
Diglossini: Highland Tanagers P.L. Sclater, 1875
- Stripe-tailed Yellow-Finch, Pseudochloris citrina
- Gray-hooded Sierra Finch, Phrygilus gayi
- Patagonian Sierra Finch, Phrygilus patagonicus
- Peruvian Sierra Finch, Phrygilus punensis
- Black-hooded Sierra Finch, Phrygilus atriceps
- Gough Finch, Rowettia goughensis
- White-bridled Finch, Melanodera melanodera
- Yellow-bridled Finch, Melanodera xanthogramma
- Inaccessible Island Finch, Nesospiza acunhae
- Nightingale Island Finch, Nesospiza questi
- Wilkins's Finch, Nesospiza wilkinsi
- Saffron Finch, Sicalis flaveola
- Grassland Yellow-Finch, Sicalis luteola
- Orange-fronted Yellow-Finch, Sicalis columbiana
- Sulphur-throated Finch, Sicalis taczanowskii
- Bright-rumped Yellow-Finch, Sicalis uropigyalis
- Citron-headed Yellow-Finch, Sicalis luteocephala
- Patagonian Yellow-Finch, Sicalis lebruni
- Greenish Yellow-Finch, Sicalis olivascens
- Monte Yellow-Finch, Sicalis mendozae
- Greater Yellow-Finch, Sicalis auriventris
- Puna Yellow-Finch, Sicalis lutea
- Raimondi's Yellow-Finch, Sicalis raimondii
- Tit-like Dacnis, Xenodacnis parina
- White-winged Diuca-Finch, Diuca speculifera
- Short-tailed Finch, Diuca brachyura
- White-throated Sierra Finch, Diuca erythronota
- Red-backed Sierra Finch, Diuca dorsalis
- Plumbeous Sierra Finch, Geospizopsis unicolor
- Ash-breasted Sierra Finch, Geospizopsis plebejus
- Uniform Finch, Haplospiza unicolor
- Slaty Finch, Haplospiza rustica
- Peg-billed Finch, Haplospiza bairdi
- Band-tailed Seedeater, Catamenia analis
- Paramo Seedeater, Catamenia homochroa
- Plain-colored Seedeater, Catamenia inornata
- Deep-blue Flower-piercer / Golden-eyed Flower-piercer, Diglossa glauca
- Bluish Flower-piercer, Diglossa caerulescens
- Masked Flower-piercer, Diglossa cyanea
- Indigo Flower-piercer, Diglossa indigotica
- Rusty Flower-piercer, Diglossa sittoides
- Cinnamon-bellied Flower-piercer, Diglossa baritula
- Slaty Flower-piercer, Diglossa plumbea
- Moustached Flower-piercer, Diglossa mystacalis
- Chestnut-bellied Flower-piercer, Diglossa gloriosissima
- Glossy Flower-piercer, Diglossa lafresnayii
- Scaled Flower-piercer, Diglossa duidae
- Greater Flower-piercer, Diglossa major
- White-sided Flower-piercer, Diglossa albilatera
- Venezuelan Flower-piercer, Diglossa venezuelensis
- Black-throated Flower-piercer, Diglossa brunneiventris
- Gray-bellied Flower-piercer, Diglossa carbonaria
- Black Flower-piercer, Diglossa humeralis
- Merida Flower-piercer, Diglossa gloriosa
Hemithraupini: Yellow-and-black Tanagers Sundevall, 1872
- Green Honeycreeper, Chlorophanes spiza
- Golden-collared Honeycreeper, Iridophanes pulcherrimus
- Black-and-yellow Tanager, Chrysothlypis chrysomelas
- Scarlet-and-white Tanager, Chrysothlypis salmoni
- Sulphur-rumped Tanager, Heterospingus rubrifrons
- Scarlet-browed Tanager, Heterospingus xanthopygius
- Yellow-backed Tanager, Hemithraupis flavicollis
- Guira Tanager, Hemithraupis guira
- Rufous-headed Tanager, Hemithraupis ruficapilla
Tachyphonini: Ornamented Tanagers Bonaparte, 1853
- Rufous-crested Tanager, Creurgops verticalis
- Slaty Tanager, Creurgops dentatus
- Blue-black Grassquit, Volatinia jacarina
- Black-and-white Tanager, Conothraupis speculigera
- Cone-billed Tanager, Conothraupis mesoleuca
- Gray-headed Tanager, Trichothraupis penicillata
- Black-goggled Tanager, Trichothraupis melanops
- Flame-crested Tanager, Trichothraupis cristata
- White-shouldered Tanager, Trichothraupis luctuosa
- Yellow-crested Tanager, Trichothraupis rufiventer
- Pileated Finch / Gray Pileated-Finch, Coryphospingus pileatus
- Red-crested Finch / Red Pileated-Finch, Coryphospingus cucullatus
- Fulvous-crested Tanager, Lanio surinamus
- Tawny-crested Tanager, Lanio delatrii
- Crimson-breasted Finch, Lanio cruentus
- White-winged Shrike-Tanager, Lanio versicolor
- Fulvous Shrike-Tanager, Lanio fulvus
- Black-throated Shrike-Tanager, Lanio aurantius
- White-throated Shrike-Tanager, Lanio leucothorax
- Red-shouldered Tanager, Tachyphonus phoenicius
- Ruby-crowned Tanager, Tachyphonus coronatus
- White-lined Tanager, Tachyphonus rufus
- Crimson-collared Tanager, Ramphocelus sanguinolentus
- Flame-rumped Tanager, Ramphocelus flammigerus
- Passerini's Tanager, Ramphocelus passerinii
- Cherrie's Tanager, Ramphocelus costaricensis
- Brazilian Tanager, Ramphocelus bresilia
- Crimson-backed Tanager, Ramphocelus dimidiatus
- Masked Crimson Tanager, Ramphocelus nigrogularis
- Silver-beaked Tanager, Ramphocelus carbo
- Black-bellied Tanager / Huallaga Tanager, Ramphocelus melanogaster
Charitospizini: Coal-crested Finch Burns et al., 2014
- Coal-crested Finch, Charitospiza eucosma
Click for Dacninae tree
part II: Charitospizini —
Poospizini
Dacnini: Blue Tanagers Sundevall, 1836
- Swallow Tanager, Tersina viridis
- Red-legged Honeycreeper, Cyanerpes cyaneus
- Purple Honeycreeper, Cyanerpes caeruleus
- Shining Honeycreeper, Cyanerpes lucidus
- Short-billed Honeycreeper, Cyanerpes nitidus
- Scarlet-thighed Dacnis, Dacnis venusta
- White-bellied Dacnis, Dacnis albiventris
- Black-legged Dacnis, Dacnis nigripes
- Blue Dacnis, Dacnis cayana
- Yellow-bellied Dacnis, Dacnis flaviventer
- Turquoise Dacnis, Dacnis hartlaubi
- Viridian Dacnis, Dacnis viguieri
- Yellow-tufted Dacnis, Dacnis egregia
- Black-faced Dacnis, Dacnis lineata
- Scarlet-breasted Dacnis, Dacnis berlepschi
Emberizoidini: Grassland Tanagers Burns et al., 2014
- Black-masked Finch, Coryphaspiza melanotis
- Great Pampa-Finch / Pampa Finch, Embernagra platensis
- Pale-throated Pampa-Finch / Serra Finch, Embernagra longicauda
- Lesser Grass-Finch, Emberizoides ypiranganus
- Wedge-tailed Grass-Finch, Emberizoides herbicola
- Duida Grass-Finch, Emberizoides duidae
Saltatorini: Saltators Bonaparte, 1853
- Many-colored Chaco Finch, Saltatricula multicolor
- Black-throated Saltator, Saltatricula atricollis
- Orinocan Saltator / Orinoco Saltator, Saltator orenocensis
- Middle American Saltator, Saltator grandis
- Plumbeous Saltator, Saltator plumbeus
- Green-winged Saltator, Saltator similis
- Grayish Saltator, Saltator coerulescens
- Streaked Saltator, Saltator striatipectus
- Lesser Antillean Saltator, Saltator albicollis
- Buff-throated Saltator, Saltator maximus
- Black-headed Saltator, Saltator atriceps
- Black-winged Saltator, Saltator atripennis
- Black-cowled Saltator, Saltator nigriceps
- Slate-colored Grosbeak, Saltator grossus
- Black-throated Grosbeak, Saltator fuliginosus
- Masked Saltator, Saltator cinctus
- Golden-billed Saltator, Saltator aurantiirostris
- Thick-billed Saltator, Saltator maxillosus
Poospizini: Warbler-Tanagers Wolters, 1980
- Cinereous Finch, Piezorina cinerea
- Slender-billed Finch, Xenospingus concolor
- Gray-hooded Bush Tanager, Cnemoscopus rubrirostris
- Black-headed Hemispingus, Pseudospingus verticalis
- Drab Hemispingus, Pseudospingus xanthophthalmus
- Bolivian Warbling-Finch, Poospiza boliviana
- Cinnamon Warbling-Finch, Poospiza ornata
- Black-and-chestnut Warbling-Finch, Poospiza whitii
- Black-and-rufous Warbling-Finch, Poospiza nigrorufa
- Collared Warbling-Finch, Poospiza hispaniolensis
- Rufous-breasted Warbling-Finch, Poospiza rubecula
- Cochabamba Mountain-Finch, Poospiza garleppi
- Tucuman Mountain-Finch, Poospiza baeri
- Slaty-backed Hemispingus, Poospiza goeringi
- Rufous-browed Hemispingus, Poospiza rufosuperciliaris
- Gray-capped Hemispingus, Kleinothraupis reyi
- Black-capped Hemispingus, Kleinothraupis atropileus
- White-browed Hemispingus, Kleinothraupis auricularis
- Parodi's Hemispingus, Kleinothraupis parodii
- Orange-browed Hemispingus, Kleinothraupis calophrys
- Piura Hemispingus, Sphenopsis piurae
- Oleaginous Hemispingus, Sphenopsis frontalis
- Black-eared Hemispingus, Sphenopsis melanotis
- Orange-headed Tanager, Thlypopsis sordida
- Buff-bellied Tanager, Thlypopsis inornata
- Fulvous-headed Tanager, Thlypopsis fulviceps
- Chestnut-headed Tanager, Thlypopsis pyrrhocoma
- Rust-and-yellow Tanager, Thlypopsis ruficeps
- Superciliaried Hemispingus, Thlypopsis superciliaris
- Rufous-chested Tanager, Thlypopsis ornata
- Brown-flanked Tanager, Thlypopsis pectoralis
- Chestnut-breasted Mountain-Finch, Poospizopsis caesar
- Rufous-sided Warbling-Finch, Poospizopsis hypocondria
- White-rumped Tanager, Cypsnagra hirundinacea
- Long-tailed Reed Finch, Donacospiza albifrons
- Bay-chested Warbling-Finch, Castanozoster thoracicus
- Black-backed Bush Tanager, Urothraupis stolzmanni
- Pardusco, Nephelornis oneilli
- Buff-throated Warbling-Finch, Microspingus lateralis
- Gray-throated Warbling-Finch, Microspingus cabanisi
- Rusty-browed Warbling-Finch, Microspingus erythrophrys
- Plain-tailed Warbling-Finch, Microspingus alticola
- Ringed Warbling-Finch, Microspingus torquatus
- Three-striped Hemispingus, Microspingus trifasciatus
- Black-capped Warbling-Finch, Microspingus melanoleucus
- Cinereous Warbling-Finch, Microspingus cinereus
Coerebini: Dome-Nesting Tanagers d'Orbigny & Lafresnaye, 1838
- Bananaquit, Coereba flaveola
Click for Dacninae tree
part III: Coerebini & Sporophilini - Yellow-faced Grassquit, Tiaris olivaceus
- Orangequit, Euneornis campestris
- Puerto Rican Bullfinch, Melopyrrha portoricensis
- Greater Antillean Bullfinch, Melopyrrha violacea
- Cuban Bullfinch, Melopyrrha nigra
- Grand Cayman Bullfinch, Melopyrrha taylori
- Yellow-shouldered Grassquit, Loxipasser anoxanthus
- Cuban Grassquit, Phonipara canora
- Lesser Antillean Bullfinch, Loxigilla noctis
- Barbados Bullfinch, Loxigilla barbadensis
- St. Lucia Black Finch, Melanospiza richardsoni
- Black-faced Grassquit, Melanospiza bicolor
- Dull-colored Grassquit, Asemospiza obscura
- Sooty Grassquit, Asemospiza fuliginosa
- Gray Warbler-Finch, Certhidea fusca
- Green Warbler-Finch, Certhidea olivacea
- Vegetarian Finch, Platyspiza crassirostris
- Cocos Finch, Pinaroloxias inornata
- Sharpe's Ground Finch, Geospiza difficilis
- Large Tree Finch, Geospiza psittacula
- Medium Tree Finch, Geospiza pauper
- Small Tree Finch, Geospiza parvula
- Mangrove Finch, Geospiza heliobates
- Woodpecker Finch, Geospiza pallida
- Vampire Finch, Geospiza septentrionalis
- Small Ground Finch, Geospiza fuliginosa
- Medium Ground Finch, Geospiza fortis
- Sharp-beaked Ground Finch, Geospiza acutirostris
- Large Ground Finch, Geospiza magnirostris
- Large-billed Cactus Finch, Geospiza conirostris
- Common Cactus Finch, Geospiza scandens
- Genovesa Cactus Finch, Geospiza propinqua
Sporophilini: Seedeaters Ridgway, 1901 (1853)
- Lesson's Seedeater, Sporophila bouvronides
- Lined Seedeater, Sporophila lineola
- White-bellied Seedeater, Sporophila leucoptera
- Parrot-billed Seedeater, Sporophila peruviana
- Chestnut-throated Seedeater, Sporophila telasco
- Drab Seedeater, Sporophila simplex
- Chestnut-bellied Seedeater, Sporophila castaneiventris
- Ruddy-breasted Seedeater, Sporophila minuta
- Black-bellied Seedeater, Sporophila melanogaster
- Copper Seedeater, Sporophila bouvreuil
- Black-and-tawny Seedeater, Sporophila nigrorufa
- Chestnut Seedeater, Sporophila cinnamomea
- Rufous-rumped Seedeater, Sporophila hypochroma
- Pearly-bellied Seedeater, Sporophila pileata
- Tawny-bellied Seedeater, Sporophila hypoxantha
- Marsh Seedeater, Sporophila palustris
- Dark-throated Seedeater, Sporophila ruficollis
- Cinnamon-rumped Seedeater, Sporophila torqueola
- Variable Seedeater, Sporophila corvina
- Gray Seedeater, Sporophila intermedia
- Caqueta Seedeater, Sporophila murallae
- Wing-barred Seedeater, Sporophila americana
- Thick-billed Seed-Finch, Sporophila funerea
- Chestnut-bellied Seed-Finch, Sporophila angolensis
- Nicaraguan Seed-Finch, Sporophila nuttingi
- Great-billed Seed-Finch, Sporophila maximiliani
- Large-billed Seed-Finch, Sporophila crassirostris
- Black-billed Seed-Finch, Sporophila atrirostris
- Morelet's Seedeater / White-collared Seedeater, Sporophila morelleti
- White-naped Seedeater, Sporophila fringilloides
- Black-and-white Seedeater, Sporophila luctuosa
- Double-collared Seedeater, Sporophila caerulescens
- Yellow-bellied Seedeater, Sporophila nigricollis
- Dubois's Seedeater, Sporophila ardesiaca
- Slate-colored Seedeater, Sporophila schistacea
- Buffy-fronted Seedeater, Sporophila frontalis
- Temminck's Seedeater, Sporophila falcirostris
- Plumbeous Seedeater, Sporophila plumbea
- Tropeiro Seedeater, Sporophila beltoni
- Rusty-collared Seedeater, Sporophila collaris
- White-throated Seedeater, Sporophila albogularis